

A role for the beta isoform of protein kinase C in fear conditioning
- PMID: 10934237
- PMCID: PMC6772611
- DOI: 10.1523/JNEUROSCI.20-16-05906.2000
A role for the beta isoform of protein kinase C in fear conditioning
Abstract
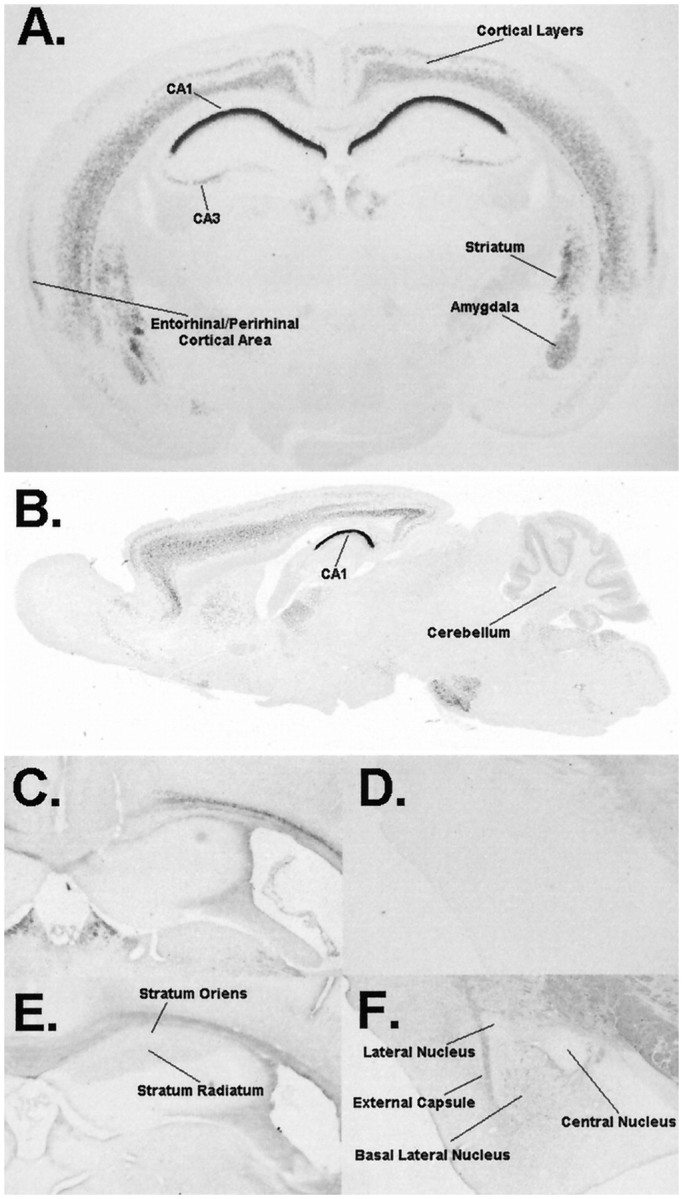
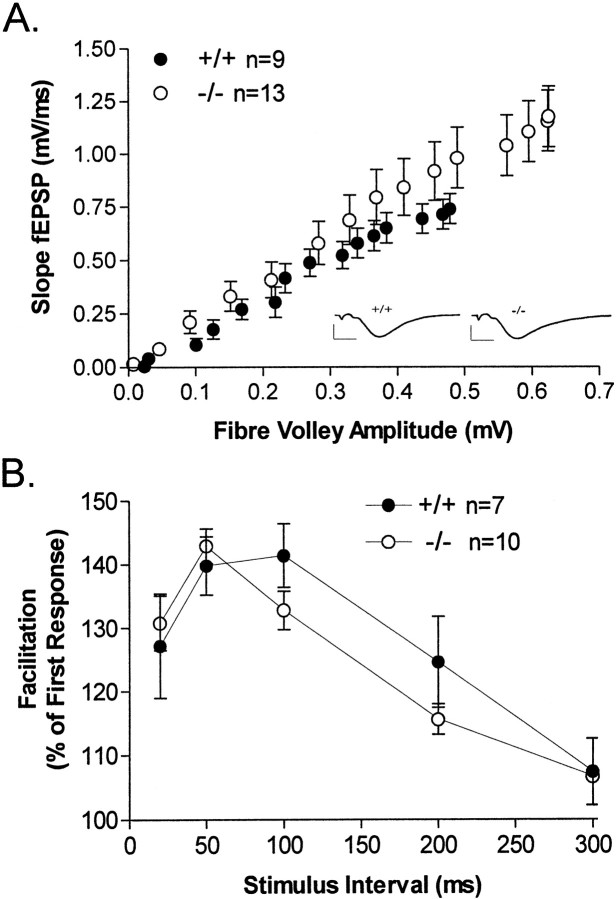
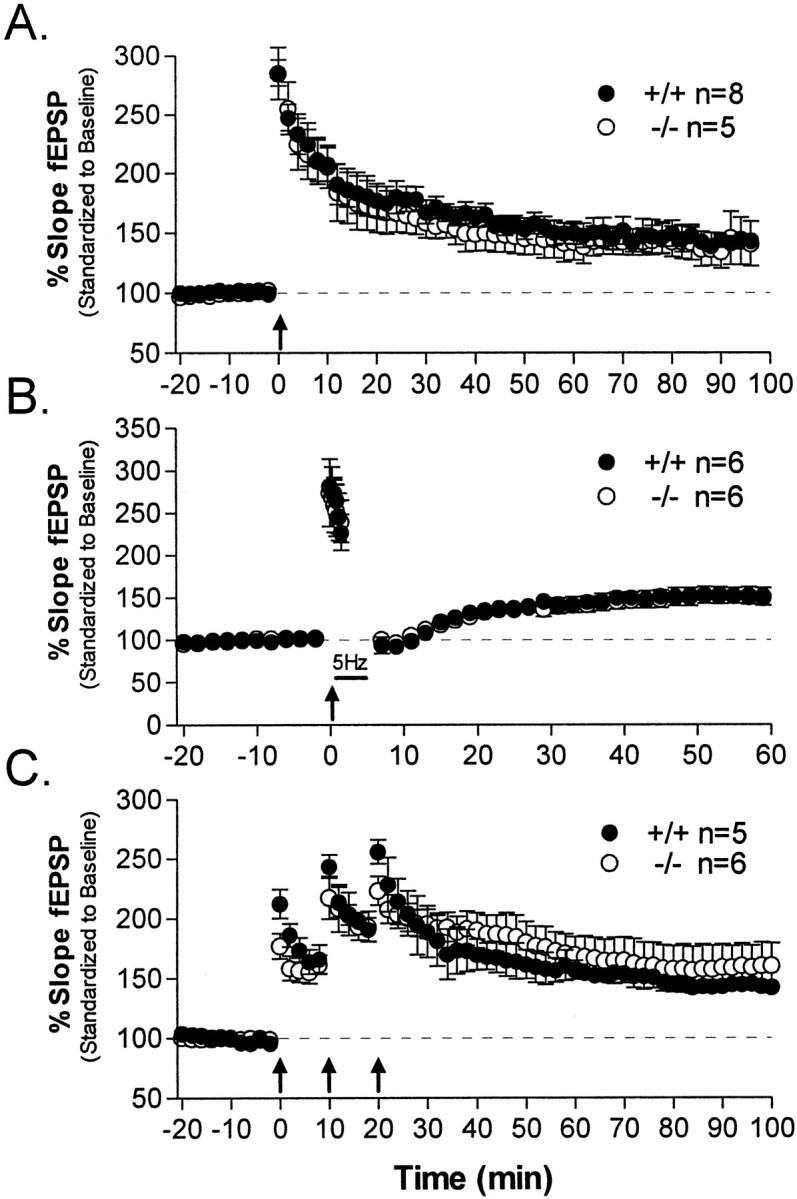
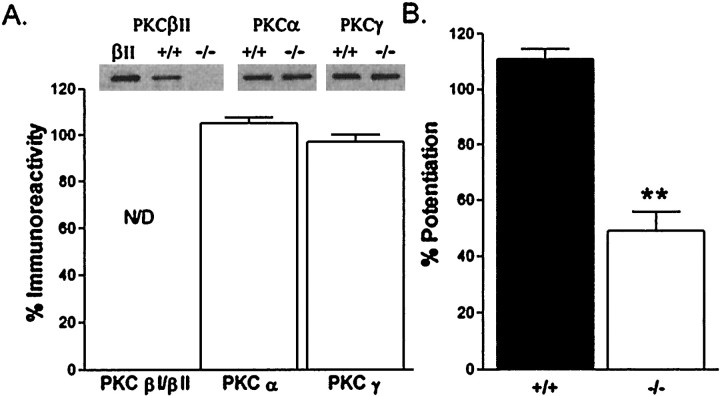
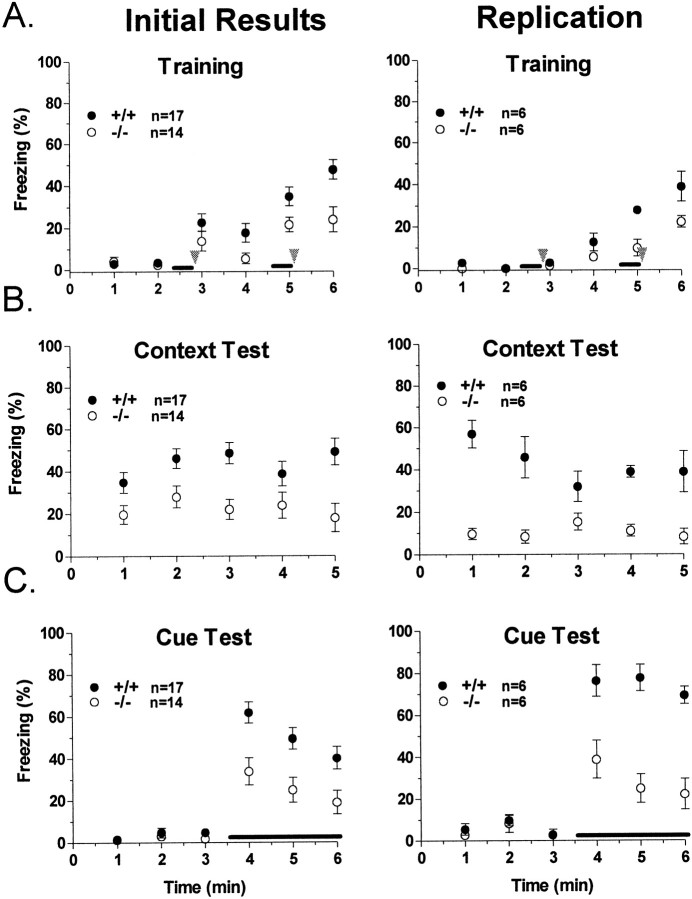
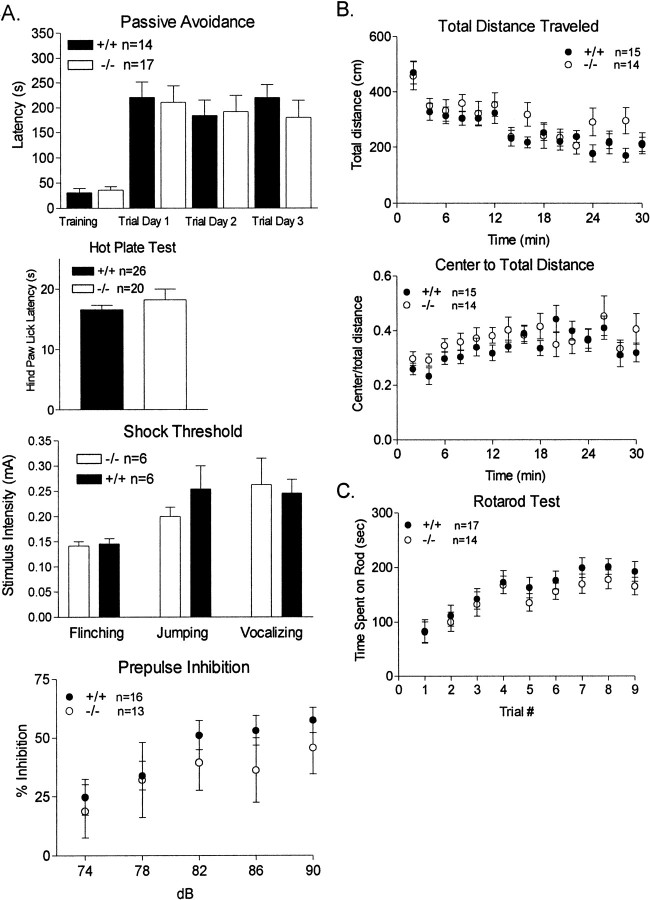
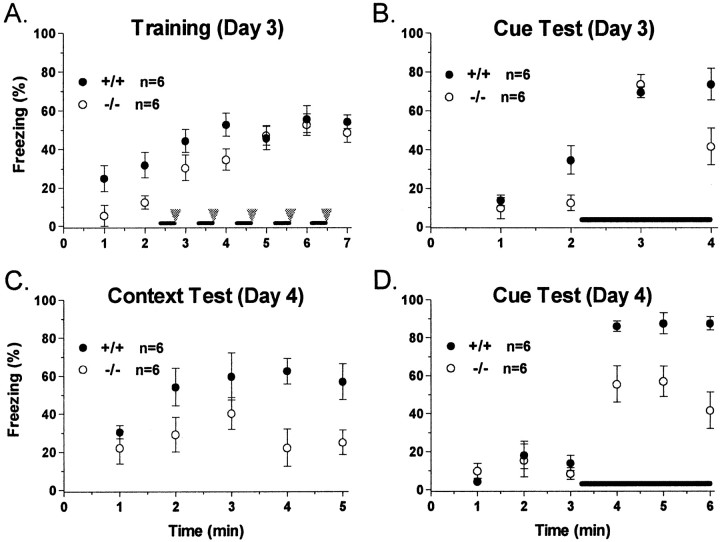
The protein kinase C family of enzymes has been implicated in synaptic plasticity and memory in a wide range of animal species, but to date little information has been available concerning specific roles for individual isoforms of this category of kinases. To investigate the role of the beta isoform of PKC in mammalian learning, we characterized mice deficient in the PKC beta gene using anatomical, biochemical, physiological, and behavioral approaches. In our studies we observed that PKC beta was predominantly expressed in the neocortex, in area CA1 of the hippocampus, and in the basolateral nucleus of the amygdala. Mice deficient in PKC beta showed normal brain anatomy and normal hippocampal synaptic transmission, paired pulse facilitation, and long-term potentiation and normal sensory and motor responses. The PKC beta knock-out animals exhibited a loss of learning, however; they suffered deficits in both cued and contextual fear conditioning. The PKC expression pattern and behavioral phenotype in the PKC beta knock-out animals indicate a critical role for the beta isoform of PKC in learning-related signal transduction mechanisms, potentially in the basolateral nucleus of the amygdala.
Figures
Similar articles
-
PKM-ζ is not required for hippocampal synaptic plasticity, learning and memory.Nature. 2013 Jan 17;493(7432):420-3. doi: 10.1038/nature11802. Epub 2013 Jan 2. Nature. 2013. PMID: 23283174 Free PMC article.
-
PKC gamma mutant mice exhibit mild deficits in spatial and contextual learning.Cell. 1993 Dec 31;75(7):1263-71. doi: 10.1016/0092-8674(93)90614-v. Cell. 1993. PMID: 8269510
-
Enhanced long-term potentiation and impaired learning in phosphodiesterase 4D-knockout (PDE4D) mice.Eur J Neurosci. 2008 Aug;28(3):625-32. doi: 10.1111/j.1460-9568.2008.06349.x. Eur J Neurosci. 2008. PMID: 18702734
-
Amygdala, long-term potentiation, and fear conditioning.Neuroscientist. 2005 Feb;11(1):75-88. doi: 10.1177/1073858404270857. Neuroscientist. 2005. PMID: 15632280 Review.
-
The neuronal MAP kinase cascade: a biochemical signal integration system subserving synaptic plasticity and memory.J Neurochem. 2001 Jan;76(1):1-10. doi: 10.1046/j.1471-4159.2001.00054.x. J Neurochem. 2001. PMID: 11145972 Review.
Cited by
-
PKCα integrates spatiotemporally distinct Ca2+ and autocrine BDNF signaling to facilitate synaptic plasticity.Nat Neurosci. 2018 Aug;21(8):1027-1037. doi: 10.1038/s41593-018-0184-3. Epub 2018 Jul 16. Nat Neurosci. 2018. PMID: 30013171 Free PMC article.
-
M1-muscarinic receptors promote fear memory consolidation via phospholipase C and the M-current.J Neurosci. 2014 Jan 29;34(5):1570-8. doi: 10.1523/JNEUROSCI.1040-13.2014. J Neurosci. 2014. PMID: 24478341 Free PMC article.
-
Early activation of extracellular signal-regulated kinase signaling pathway in the hippocampus is required for short-term memory formation of a fear-motivated learning.Cell Mol Neurobiol. 2006 Jul-Aug;26(4-6):989-1002. doi: 10.1007/s10571-006-9116-y. Cell Mol Neurobiol. 2006. PMID: 16977492 Free PMC article.
-
Learning and memory deficits in mice lacking protease activated receptor-1.Neurobiol Learn Mem. 2007 Oct;88(3):295-304. doi: 10.1016/j.nlm.2007.04.004. Epub 2007 Jun 1. Neurobiol Learn Mem. 2007. PMID: 17544303 Free PMC article.
-
Molecular mechanisms of fear learning and memory.Cell. 2011 Oct 28;147(3):509-24. doi: 10.1016/j.cell.2011.10.009. Cell. 2011. PMID: 22036561 Free PMC article. Review.
References
-
- Abel T, Nguyen PV, Barad M, Deuel TA, Kandel ER, Bourtchouladze R. Genetic demonstration of a role for PKA in the late phase of LTP and in hippocampus-based long-term memory. Cell. 1997;88:615–626. - PubMed
-
- Abeliovich A, Paylor R, Chen C, Kim JJ, Wehner JM, Tonegawa S. PKC gamma mutant mice exhibit mild deficits in spatial and contextual learning. Cell. 1993;75:1263–1271. - PubMed
-
- Bordi F, Marcon C, Chiamulera C, Reggiani A. Effects of the metabotropic glutamate receptor antagonist MCPG on spatial and context-specific learning. Neuropharmacology. 1996;35:1557–1565. - PubMed
-
- Chen SJ, Sweatt JD, Klann E. Enhanced phosphorylation of the postsynaptic protein kinase C substrate RC3/neurogranin during long-term potentiation. Brain Res. 1997;749:181–187. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous