

The Rad51 paralog Rad51B promotes homologous recombinational repair
- PMID: 10938124
- PMCID: PMC86122
- DOI: 10.1128/MCB.20.17.6476-6482.2000
The Rad51 paralog Rad51B promotes homologous recombinational repair
Abstract
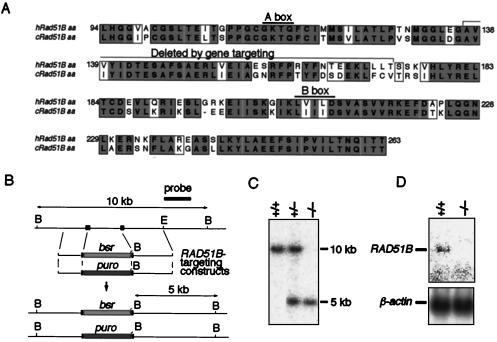
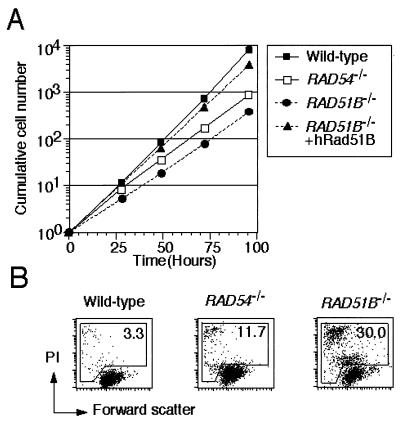
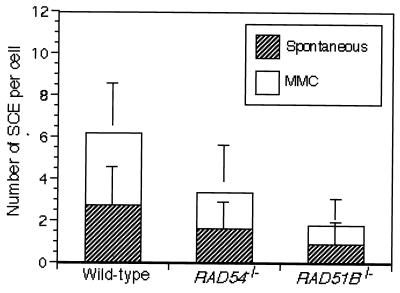
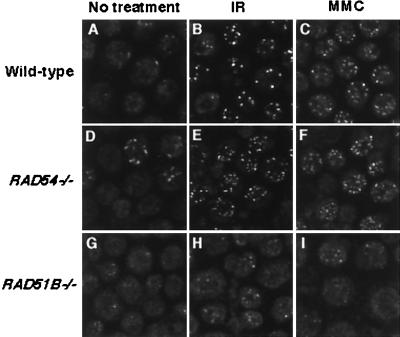
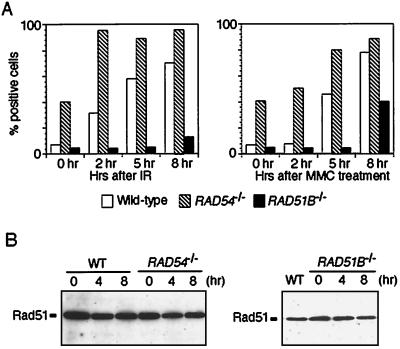
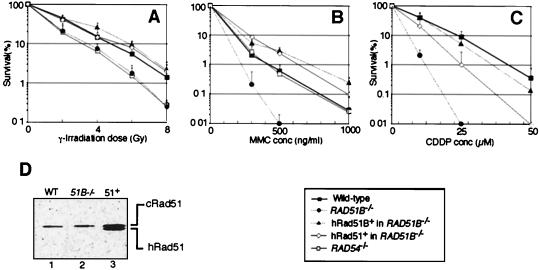
The highly conserved Saccharomyces cerevisiae Rad51 protein plays a central role in both mitotic and meiotic homologous DNA recombination. Seven members of the Rad51 family have been identified in vertebrate cells, including Rad51, Dmc1, and five Rad51-related proteins referred to as Rad51 paralogs, which share 20 to 30% sequence identity with Rad51. In chicken B lymphocyte DT40 cells, we generated a mutant with RAD51B/RAD51L1, a member of the Rad51 family, knocked out. RAD51B(-/-) cells are viable, although spontaneous chromosomal aberrations kill about 20% of the cells in each cell cycle. Rad51B deficiency impairs homologous recombinational repair (HRR), as measured by targeted integration, sister chromatid exchange, and intragenic recombination at the immunoglobulin locus. RAD51B(-/-) cells are quite sensitive to the cross-linking agents cisplatin and mitomycin C and mildly sensitive to gamma-rays. The formation of damage-induced Rad51 nuclear foci is much reduced in RAD51B(-/-) cells, suggesting that Rad51B promotes the assembly of Rad51 nucleoprotein filaments during HRR. These findings show that Rad51B is important for repairing various types of DNA lesions and maintaining chromosome integrity.
Figures
References
-
- Albala J S, Thelen M P, Prange C, Fan W, Christensen M, Thompson L H, Lennon G G. Identification of a novel human RAD51 homolog, RAD51B. Genomics. 1997;46:476–479. - PubMed
-
- Baumann P, West S C. Role of the human RAD51 protein in homologous recombination and double-stranded-break repair. Trends Biochem Sci. 1998;23:247–251. - PubMed
-
- Bezzubova O Y, Silbergleit A, Yamaguchi-Iwai Y, Takeda S, Buerstedde J M. Reduced X-ray resistance and homologous recombination frequencies in a RAD54−/− mutant of the chicken DT40 cell line. Cell. 1997;89:185–193. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials