

Phosphoenolpyruvate carboxykinase is necessary for the integration of hepatic energy metabolism
- PMID: 10938127
- PMCID: PMC86125
- DOI: 10.1128/MCB.20.17.6508-6517.2000
Phosphoenolpyruvate carboxykinase is necessary for the integration of hepatic energy metabolism
Abstract
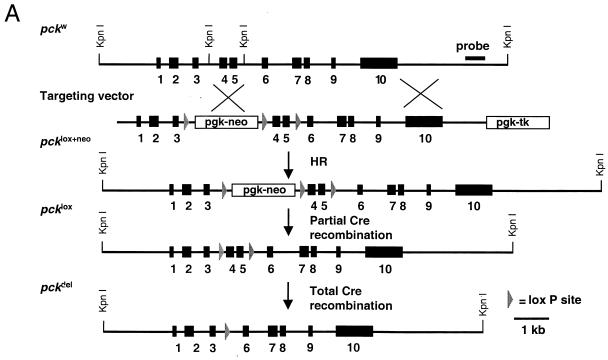
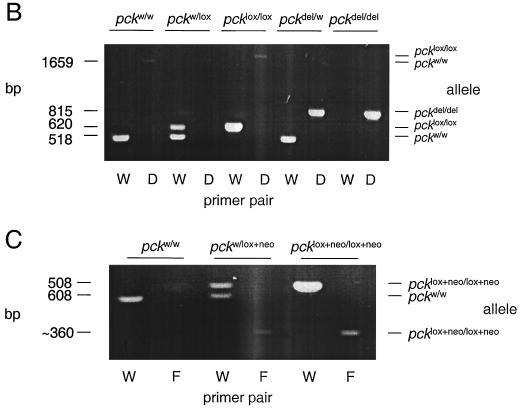
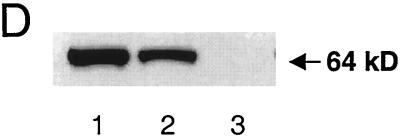
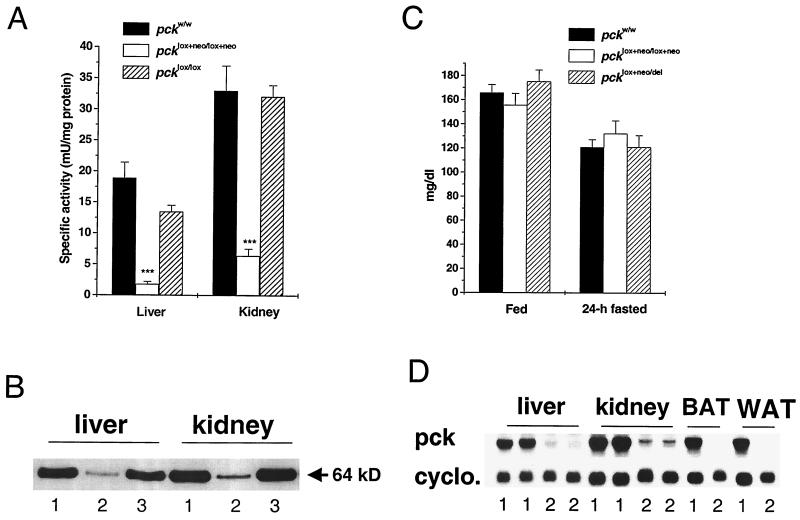
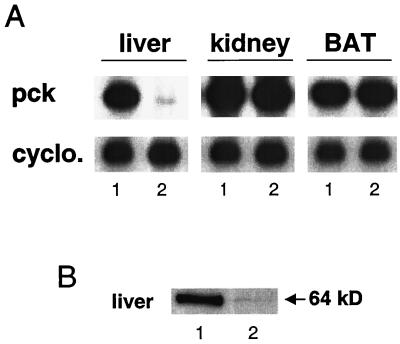
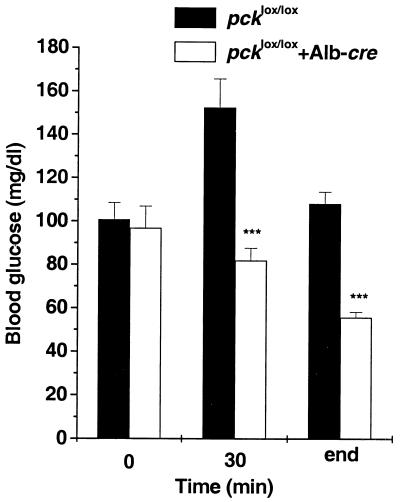
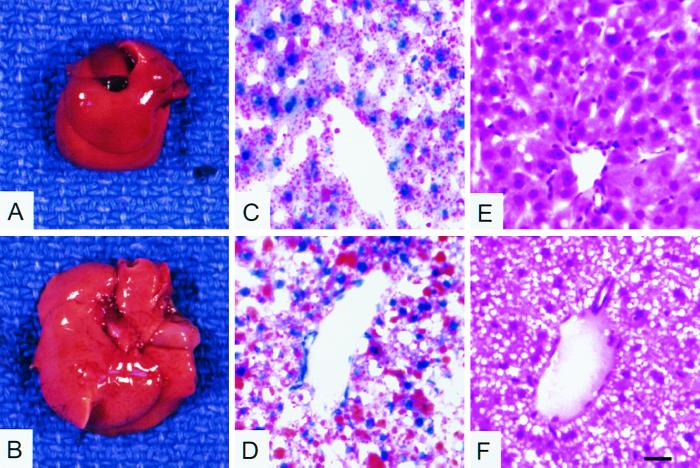
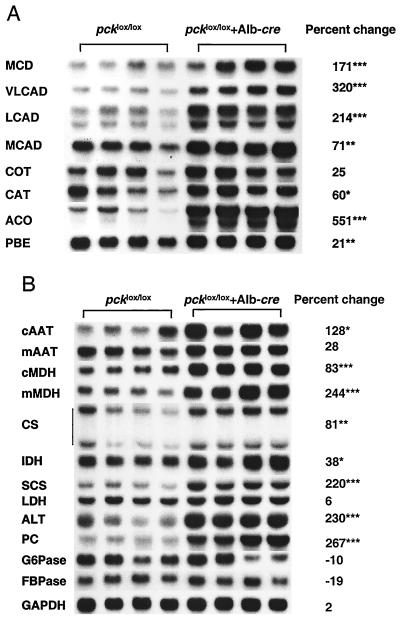
We used an allelogenic Cre/loxP gene targeting strategy in mice to determine the role of cytosolic phosphoenolpyruvate carboxykinase (PEPCK) in hepatic energy metabolism. Mice that lack this enzyme die within 3 days of birth, while mice with at least a 90% global reduction of PEPCK, or a liver-specific knockout of PEPCK, are viable. Surprisingly, in both cases these animals remain euglycemic after a 24-h fast. However, mice without hepatic PEPCK develop hepatic steatosis after fasting despite up-regulation of a variety of genes encoding free fatty acid-oxidizing enzymes. Also, marked alterations in the expression of hepatic genes involved in energy metabolism occur in the absence of any changes in plasma hormone concentrations. Given that a ninefold elevation of the hepatic malate concentration occurs in the liver-specific PEPCK knockout mice, we suggest that one or more intermediary metabolites may directly regulate expression of the affected genes. Thus, hepatic PEPCK may function more as an integrator of hepatic energy metabolism than as a determinant of gluconeogenesis.
Figures
Similar articles
-
Cytosolic phosphoenolpyruvate carboxykinase does not solely control the rate of hepatic gluconeogenesis in the intact mouse liver.Cell Metab. 2007 Apr;5(4):313-20. doi: 10.1016/j.cmet.2007.03.004. Cell Metab. 2007. PMID: 17403375 Free PMC article.
-
PEPCK-M expression in mouse liver potentiates, not replaces, PEPCK-C mediated gluconeogenesis.J Hepatol. 2013 Jul;59(1):105-13. doi: 10.1016/j.jhep.2013.02.020. Epub 2013 Mar 4. J Hepatol. 2013. PMID: 23466304 Free PMC article.
-
Vitamin A regulates genes involved in hepatic gluconeogenesis in mice: phosphoenolpyruvate carboxykinase, fructose-1,6-bisphosphatase and 6-phosphofructo-2-kinase/fructose-2,6-bisphosphatase.J Nutr. 1997 Jul;127(7):1274-8. doi: 10.1093/jn/127.7.1274. J Nutr. 1997. PMID: 9202079
-
Regulation of phosphoenolpyruvate carboxykinase (GTP) gene expression.Annu Rev Biochem. 1997;66:581-611. doi: 10.1146/annurev.biochem.66.1.581. Annu Rev Biochem. 1997. PMID: 9242918 Review.
-
PCK1 and PCK2 as candidate diabetes and obesity genes.Cell Biochem Biophys. 2007;48(2-3):89-95. doi: 10.1007/s12013-007-0025-6. Cell Biochem Biophys. 2007. PMID: 17709878 Review.
Cited by
-
Phosphoenolpyruvate carboxykinase and glucose-6-phosphatase are required for steroidogenesis in testicular Leydig cells.J Biol Chem. 2012 Dec 7;287(50):41875-87. doi: 10.1074/jbc.M112.421552. Epub 2012 Oct 16. J Biol Chem. 2012. PMID: 23074219 Free PMC article.
-
Suppression of diet-induced hypercholesterolemia by turtle jelly, a traditional chinese functional food, in rats.Evid Based Complement Alternat Med. 2012;2012:320304. doi: 10.1155/2012/320304. Epub 2012 Oct 31. Evid Based Complement Alternat Med. 2012. PMID: 23243438 Free PMC article.
-
Phosphoenolpyruvate carboxykinase in cucumber plants is increased both by ammonium and by acidification, and is present in the phloem.Planta. 2004 May;219(1):48-58. doi: 10.1007/s00425-004-1220-y. Epub 2004 Feb 26. Planta. 2004. PMID: 14991407
-
The mitochondrial isoform of phosphoenolpyruvate carboxykinase (PEPCK-M) and glucose homeostasis: has it been overlooked?Biochim Biophys Acta. 2014 Apr;1840(4):1313-30. doi: 10.1016/j.bbagen.2013.10.033. Epub 2013 Oct 28. Biochim Biophys Acta. 2014. PMID: 24177027 Free PMC article. Review.
-
Pyruvate-Carboxylase-Mediated Anaplerosis Promotes Antioxidant Capacity by Sustaining TCA Cycle and Redox Metabolism in Liver.Cell Metab. 2019 Jun 4;29(6):1291-1305.e8. doi: 10.1016/j.cmet.2019.03.014. Epub 2019 Apr 18. Cell Metab. 2019. PMID: 31006591 Free PMC article.
References
-
- Bergmeyer H U. and 1985. Methods of enzymatic analysis. 3rd ed. 4 to 6. Weinheim, Germany: VCH Verlagsgesllschaft; 1984.
-
- Cersosimo E, Molina P E, Abumrad N N. Renal glucose production during insulin-induced hypoglycemia. Diabetes. 1997;46:643–646. . (Erratum, 46:1532.) - PubMed
-
- Cimbala M A, Lamers W H, Nelson K, Monahan J E, Yoo-Warren H, Hanson R W. Rapid changes in the concentration of phosphoenolpyruvate carboxykinase mRNA in rat liver and kidney. Effects of insulin and cyclic AMP. J Biol Chem. 1982;257:7629–7636. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases