

Targeted inactivation of the plastid ndhB gene in tobacco results in an enhanced sensitivity of photosynthesis to moderate stomatal closure
- PMID: 10938352
- PMCID: PMC59092
- DOI: 10.1104/pp.123.4.1337
Targeted inactivation of the plastid ndhB gene in tobacco results in an enhanced sensitivity of photosynthesis to moderate stomatal closure
Abstract
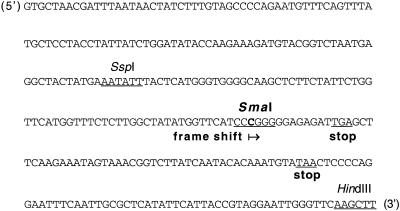
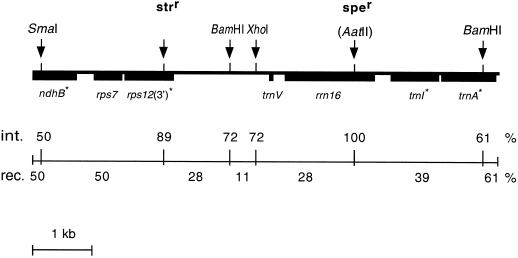
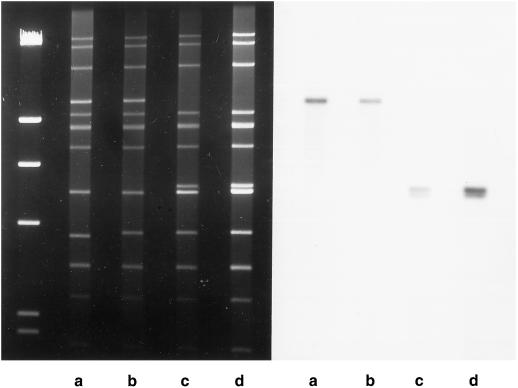
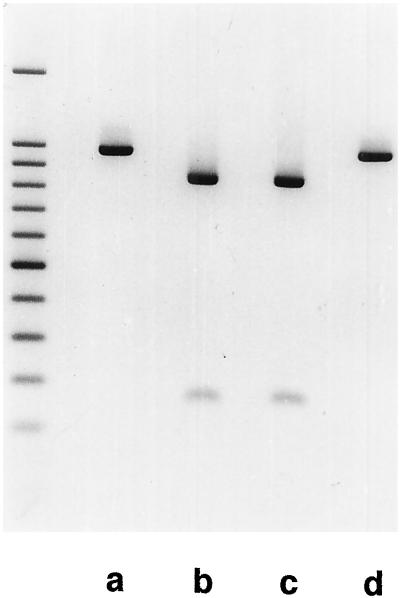
The ndh genes encoding for the subunits of NAD(P)H dehydrogenase complex represent the largest family of plastid genes without a clearly defined function. Tobacco (Nicotiana tabacum) plastid transformants were produced in which the ndhB gene was inactivated by replacing it with a mutant version possessing translational stops in the coding region. Western-blot analysis indicated that no functional NAD(P)H dehydrogenase complex can be assembled in the plastid transformants. Chlorophyll fluorescence measurements showed that dark reduction of the plastoquinone pool by stromal reductants was impaired in ndhB-inactivated plants. Both the phenotype and photosynthetic performance of the plastid transformants was completely normal under favorable conditions. However, an enhanced growth retardation of ndhB-inactivated plants was revealed under humidity stress conditions causing a moderate decline in photosynthesis via stomatal closure. This distinctive phenotype was mimicked under normal humidity by spraying plants with abscisic acid. Measurements of CO(2) fixation demonstrated an enhanced decline in photosynthesis in the mutant plants under humidity stress, which could be restored to wild-type levels by elevating the external CO(2) concentration. These results suggest that the plastid NAD(P)H:plastoquinone oxidoreductase in tobacco performs a significant physiological role by facilitating photosynthesis at moderate CO(2) limitation.
Figures


Similar articles
-
Increased sensitivity of photosynthesis to antimycin A induced by inactivation of the chloroplast ndhB gene. Evidence for a participation of the NADH-dehydrogenase complex to cyclic electron flow around photosystem I.Plant Physiol. 2001 Apr;125(4):1919-29. doi: 10.1104/pp.125.4.1919. Plant Physiol. 2001. PMID: 11299371 Free PMC article.
-
New subunits NDH-M, -N, and -O, encoded by nuclear genes, are essential for plastid Ndh complex functioning in higher plants.Plant Cell. 2005 Jan;17(1):219-32. doi: 10.1105/tpc.104.028282. Epub 2004 Dec 17. Plant Cell. 2005. PMID: 15608332 Free PMC article.
-
Expression of the plastid ndhF gene product in photosynthetic and non-photosynthetic tissues of developing barley seedlings.Plant Cell Physiol. 1997 Dec;38(12):1382-8. doi: 10.1093/oxfordjournals.pcp.a029133. Plant Cell Physiol. 1997. PMID: 9522468
-
Plastid ndh genes in plant evolution.Plant Physiol Biochem. 2010 Aug;48(8):636-45. doi: 10.1016/j.plaphy.2010.04.009. Epub 2010 May 9. Plant Physiol Biochem. 2010. PMID: 20493721 Review.
-
Chlororespiration and cyclic electron flow around PSI during photosynthesis and plant stress response.Plant Cell Environ. 2007 Sep;30(9):1041-51. doi: 10.1111/j.1365-3040.2007.01675.x. Plant Cell Environ. 2007. PMID: 17661746 Review.
Cited by
-
Strategies for adaptation to high light in plants.aBIOTECH. 2024 May 13;5(3):381-393. doi: 10.1007/s42994-024-00164-6. eCollection 2024 Sep. aBIOTECH. 2024. PMID: 39279858 Free PMC article. Review.
-
Flexibility in photosynthetic electron transport: a newly identified chloroplast oxidase involved in chlororespiration.Philos Trans R Soc Lond B Biol Sci. 2000 Oct 29;355(1402):1447-54. doi: 10.1098/rstb.2000.0705. Philos Trans R Soc Lond B Biol Sci. 2000. PMID: 11127998 Free PMC article.
-
Plastid genome evolution in Amazonian açaí palm (Euterpe oleracea Mart.) and Atlantic forest açaí palm (Euterpe edulis Mart.).Plant Mol Biol. 2021 Mar;105(4-5):559-574. doi: 10.1007/s11103-020-01109-5. Epub 2021 Jan 1. Plant Mol Biol. 2021. PMID: 33386578
-
The complete plastomes of red fleshed pitaya (Selenicereus monacanthus) and three related Selenicereus species: insights into gene losses, inverted repeat expansions and phylogenomic implications.Physiol Mol Biol Plants. 2022 Jan;28(1):123-137. doi: 10.1007/s12298-021-01121-z. Epub 2022 Jan 11. Physiol Mol Biol Plants. 2022. PMID: 35221575 Free PMC article.
-
IMMUTANS does not act as a stress-induced safety valve in the protection of the photosynthetic apparatus of Arabidopsis during steady-state photosynthesis.Plant Physiol. 2006 Oct;142(2):574-85. doi: 10.1104/pp.106.085886. Epub 2006 Aug 4. Plant Physiol. 2006. PMID: 16891546 Free PMC article.
References
-
- Arizmendi JM, Runswick MJ, Skehel JM, Walker JE. NADH:ubiquinone oxidoreductase from bovine heart mitochondria: a fourth nuclear encoded subunit with a homologue encoded in chloroplast genomes. FEBS Lett. 1992;301:237–242. - PubMed
-
- Berger S, Ellersiek U, Westhoff P, Steinmüller K. Studies on the expression of NDH-H, a subunit of the NAD(P) H-plastoquinone-oxidoreductase of higher-plant chloroplasts. Planta. 1993;190:25–31.
-
- Bookjans G, Stummann BM, Henningsen KW. Preparation of chloroplast DNA from pea plastids isolated in a medium of high ionic strength. Anal Biochem. 1984;141:244–247. - PubMed
-
- Catalá R, Sabater B, Guéra A. Expression of the plastid ndhF gene product in photosynthetic and non-photosynthetic tissues of developing barley seedlings. Plant Cell Physiol. 1997;38:1382–1388. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
