

A plastid terminal oxidase associated with carotenoid desaturation during chromoplast differentiation
- PMID: 10938359
- PMCID: PMC59099
- DOI: 10.1104/pp.123.4.1427
A plastid terminal oxidase associated with carotenoid desaturation during chromoplast differentiation
Abstract
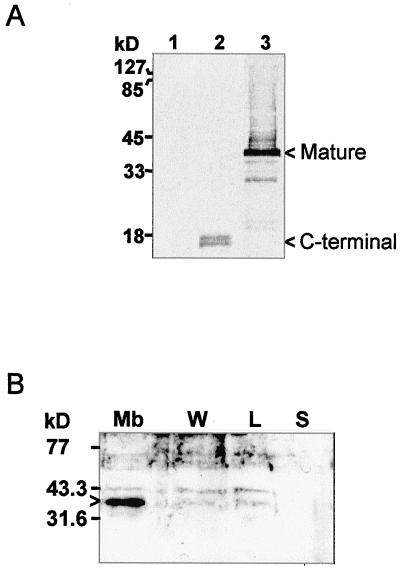
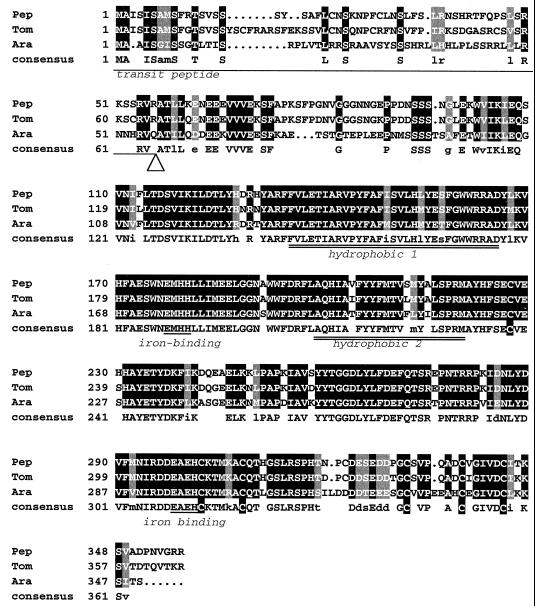
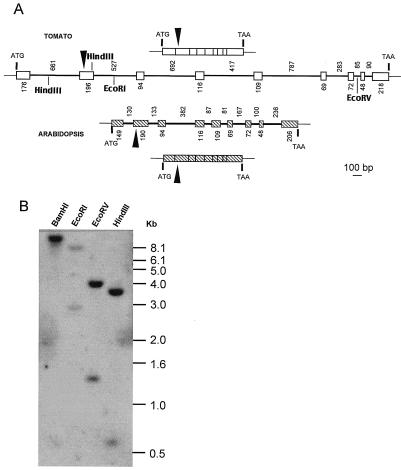
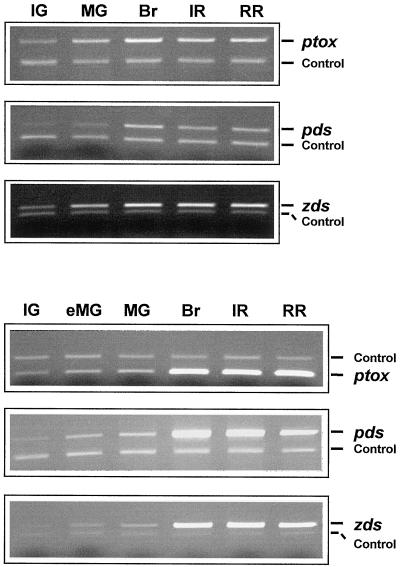
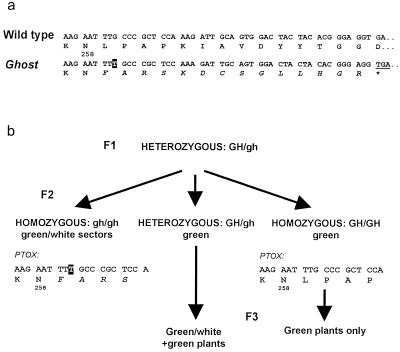
The Arabidopsis IMMUTANS gene encodes a plastid homolog of the mitochondrial alternative oxidase, which is associated with phytoene desaturation. Upon expression in Escherichia coli, this protein confers a detectable cyanide-resistant electron transport to isolated membranes. In this assay this activity is sensitive to n-propyl-gallate, an inhibitor of the alternative oxidase. This protein appears to be a plastid terminal oxidase (PTOX) that is functionally equivalent to a quinol:oxygen oxidoreductase. This protein was immunodetected in achlorophyllous pepper (Capsicum annuum) chromoplast membranes, and a corresponding cDNA was cloned from pepper and tomato (Lycopersicum esculentum) fruits. Genomic analysis suggests the presence of a single gene in these organisms, the expression of which parallels phytoene desaturase and zeta-carotene desaturase gene expression during fruit ripening. Furthermore, this PTOX gene is impaired in the tomato ghost mutant, which accumulates phytoene in leaves and fruits. These data show that PTOX also participates in carotenoid desaturation in chromoplasts in addition to its role during early chloroplast development.
Figures

References
-
- Albrecht M, Klein A, Hugueney P, Sandmann G, Kuntz M. Molecular cloning and functional expression in E. coli of a novel plant enzyme mediating ζ-carotene desaturation. FEBS Lett. 1995;372:199–202. - PubMed
-
- Andersson ME, Nordlung P. A revised model for the active site of alternative oxidase. FEBS Lett. 1999;449:17–22. - PubMed
-
- Bartley GE, Scolnik PA, Beyer P. Two Arabidopsis thaliana carotene desaturase, expressed in Escherichia coli, catalyze a poly-cis pathway to yield pro-lycopene. Eur J Biochem. 1999;259:396–403. - PubMed
-
- Berthold DA. Isolation of mutants of the Arabidopsis thaliana alternative oxidase (ubiquinol:oxygen oxidoreductase) resistant to salicylhydroxamic acid. Biochim Biophys Acta. 1998;1364:73–83. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
