

Directed evolution of a (beta alpha)8-barrel enzyme to catalyze related reactions in two different metabolic pathways
- PMID: 10944186
- PMCID: PMC27628
- DOI: 10.1073/pnas.160255397
Directed evolution of a (beta alpha)8-barrel enzyme to catalyze related reactions in two different metabolic pathways
Abstract
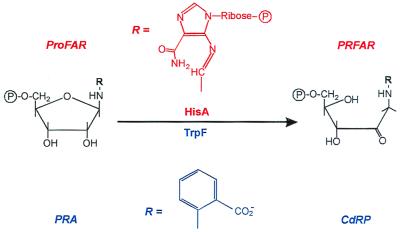
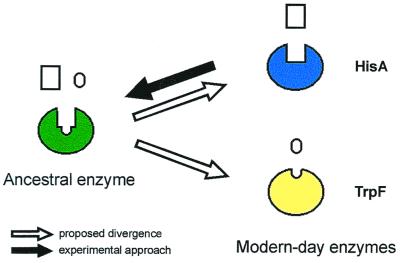
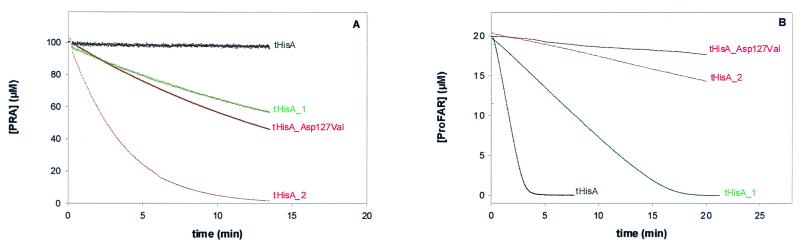
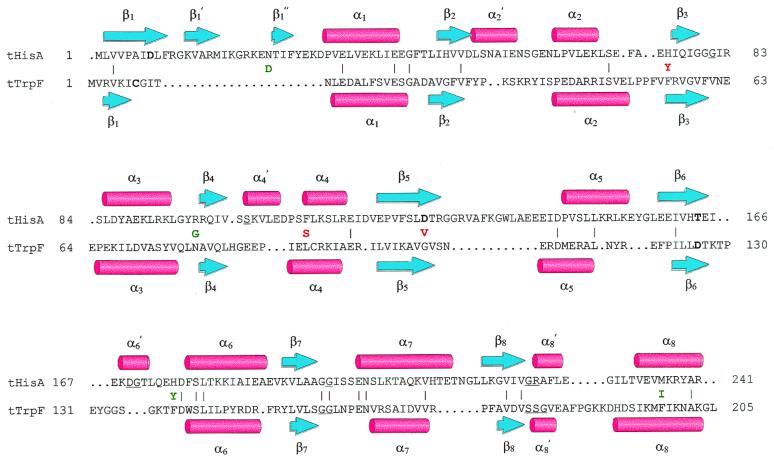
Enzymes participating in different metabolic pathways often have similar catalytic mechanisms and structures, suggesting their evolution from a common ancestral precursor enzyme. We sought to create a precursor-like enzyme for N'-[(5'-phosphoribosyl)formimino]-5-aminoimidazole-4-carboxamide ribonucleotide (ProFAR) isomerase (HisA; EC ) and phosphoribosylanthranilate (PRA) isomerase (TrpF; EC ), which catalyze similar reactions in the biosynthesis of the amino acids histidine and tryptophan and have a similar (betaalpha)(8)-barrel structure. Using random mutagenesis and selection, we generated several HisA variants that catalyze the TrpF reaction both in vivo and in vitro, and one of these variants retained significant HisA activity. A more detailed analysis revealed that a single amino acid exchange could establish TrpF activity on the HisA scaffold. These findings suggest that HisA and TrpF may have evolved from an ancestral enzyme of broader substrate specificity and underscore that (betaalpha)(8)-barrel enzymes are very suitable for the design of new catalytic activities.
Figures
References
-
- Gerlt J A, Babbitt P C. Curr Opin Chem Biol. 1998;2:607–612. - PubMed
-
- Altamirano M M, Blackburn J M, Aguayo C, Fersht A R. Nature (London) 2000;403:617–622. - PubMed
-
- Jensen R A. Annu Rev Microbiol. 1976;30:409–425. - PubMed
-
- Winkler M E. In: Escherichia coli and Salmonella: Cellular and Molecular Biology. 2nd Ed. Neidhardt F C, Ingraham J L, Low K B, Magasanik B, Schaechter M, Umbarger H E, editors. Vol. 1. Washington, DC: Am. Soc. Microbiol.; 1996. pp. 485–505.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
