

The human papillomavirus type 16 E6 and E7 oncoproteins cooperate to induce mitotic defects and genomic instability by uncoupling centrosome duplication from the cell division cycle
- PMID: 10944189
- PMCID: PMC27652
- DOI: 10.1073/pnas.170093297
The human papillomavirus type 16 E6 and E7 oncoproteins cooperate to induce mitotic defects and genomic instability by uncoupling centrosome duplication from the cell division cycle
Abstract
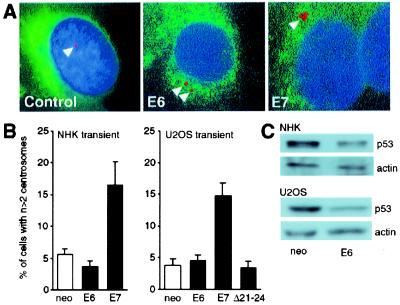
Loss of genomic integrity is a defining feature of many human malignancies, including human papillomavirus (HPV)-associated preinvasive and invasive genital squamous lesions. Here we show that aberrant mitotic spindle pole formation caused by abnormal centrosome numbers represents an important mechanism in accounting for numeric chromosomal alterations in HPV-associated carcinogenesis. Similar to what we found in histopathological specimens, HPV-16 E6 and E7 oncoproteins cooperate to induce abnormal centrosome numbers, aberrant mitotic spindle pole formation, and genomic instability. The low-risk HPV-6 E6 and E7 proteins did not induce such abnormalities. Whereas the HPV-16 E6 oncoprotein has no immediate effects on centrosome numbers, HPV-16 E7 rapidly induces abnormal centrosome duplication. Thus our results suggest a model whereby HPV-16 E7 induces centrosome-related mitotic disturbances that are potentiated by HPV-16 E6.
Figures




References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
