

doi: 10.1073/pnas.97.17.9419.
Analysis of topoisomerase function in bacterial replication fork movement: use of DNA microarrays
Affiliations
- PMID: 10944214
- PMCID: PMC16879
- DOI: 10.1073/pnas.97.17.9419
Item in Clipboard
Analysis of topoisomerase function in bacterial replication fork movement: use of DNA microarrays
Proc Natl Acad Sci U S A.
.
Abstract
We used DNA microarrays of the Escherichia coli genome to trace the progression of chromosomal replication forks in synchronized cells. We found that both DNA gyrase and topoisomerase IV (topo IV) promote replication fork progression. When both enzymes were inhibited, the replication fork stopped rapidly. The elongation rate with topo IV alone was 1/3 of normal. Genetic data confirmed and extended these results. Inactivation of gyrase alone caused a slow stop of replication. Topo IV activity was sufficient to prevent accumulation of (+) supercoils in plasmid DNA in vivo, suggesting that topo IV can promote replication by removing (+) supercoils in front of the chromosomal fork.
Figures
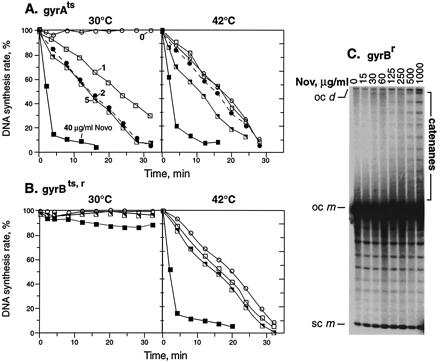
Effects on DNA replication of novobiocin addition and mutational inactivation of gyrase. Novobiocin (Nov) was added to acrAgyrA43ts cells (A) or acrA gyrBts,r novobiocin-resistant cells (B) at time 0. Cells were pulsed for 1 min with [3H]thymidine at each time point and acid precipitable counts determined. (A and B) Novobiocin concentrations were (○) 0 μg/ml; (□) 1 μg/ml; (●) 2 μg/ml; (└) 5 μg/ml; and (▪) 40 μg/ml. The rate of DNA synthesis shown is the percentage of the rate in an isogenic wild-type strain grown in parallel but without drug. (C) A acrA+gyrBr strain was treated with the indicated concentrations of novobiocin for 15 min at 30°C, and plasmid DNA from the strain was nicked and analyzed by agarose gel electrophoresis. Positions of catenanes, supercoiled monomers (sc m), open circular monomer (oc m), and open circular dimer (oc d) are indicated.
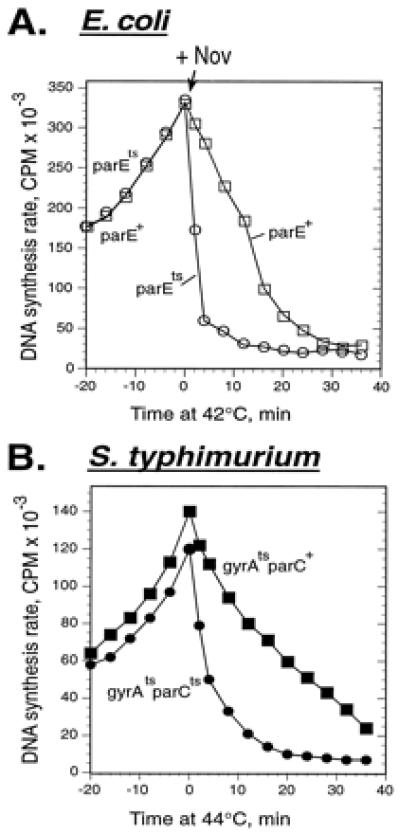
In the absence of gyrase, inhibition of topo IV causes rapid replication arrest. (A) E. coli strains acrA parE+ (□) and acrA parEts (○) were grown at 30°C. At time 0, novobiocin (Nov) was added to 5 μg/ml, and the cultures were simultaneously shifted to 42°C. The rates of DNA synthesis were measured at the indicated times. (B) S. typhimurium strains gyrA208 (gyrAts, ▪) and gyrA208parEts (●) were grown at 30°C. At time 0, they were shifted to 44°C. The rates of DNA synthesis were measured at the indicated times after temperature shift.
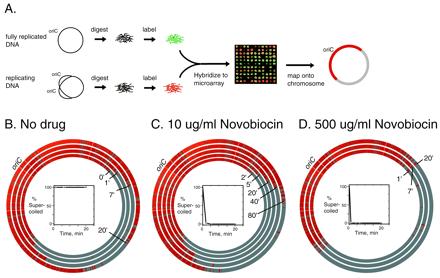
Roles of topo IV and gyrase in replication fork progression. (A) Schematic illustrating the use of genome microarrays to trace replication fork progression. Replication was synchronized in a temperature-sensitive dnaC2 initiation mutant. Fluorescently labeled genomic DNAs from a synchronized replicating culture (red) and a fully replicated reference DNA (green) were hybridized together to a glass slide containing DNA from 4,115 individual ORFs of E. coli. A representative sector of an array is shown. The fluorescence ratio in each spot indicates whether the gene was replicated. Each spot was mapped to its position on the E. coli chromosome. Replicated genes are shown in red. (B–D) Plasmid pBR322 and genomic DNAs were isolated from the cells at the indicated time points. The first time point (“0”) from the replicating DNA was taken 30 min after the shift to 30°C and immediately before the addition of novobiocin to the indicated concentrations. The positions of the clockwise replication forks at times after inhibition of gyrase or of gyrase and topo IV are shown as concentric circles superimposed on the E. coli physical map. oriC marks the origin of replication. The percent of pBR322 DNA that was supercoiled at each time point was determined by agarose gel electrophoresis and is shown in the Insets.
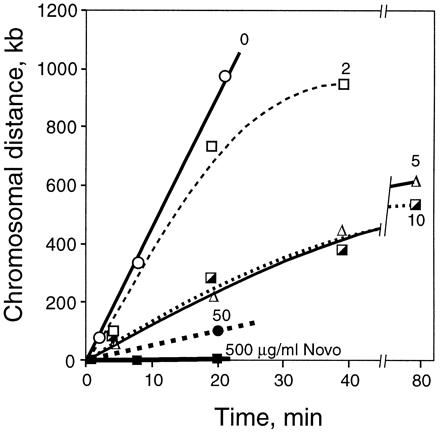
Rates of chromosomal replication supported by gyrase and topo IV. acrA dnaC2 cells were synchronized, and the position of the replication forks was determined by using hybridization to a genomic microarray as shown in Fig. 3. The plots show the average distance of the two forks relative to their respective positions at the time of novobiocin addition (time 0). Thirty minutes after the shift to 30°C to restart replication, novobiocin was added to 0 (○), 2 (□), 5 (Δ), 10 (┌), 50 (●), or 500 (▪) μg/ml. The times plotted are after addition of drug. The initial slopes (between 0 and 20 min) of the curves are: 0 drug, 45 kb/min; 2 μg/ml novobiocin, 35 kb/min; 5 and 10 μg/ml, 13.5 kb/min; 50 μg/ml, 5.1 kb/min; and 500 μg/ml, 0 kb/min.
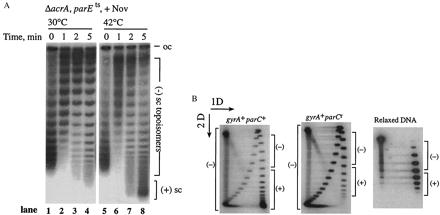
Topo IV efficiently relaxes positively supercoiled DNA in vivo. (A) pBR322 DNA was isolated from an E. coli ΔacrAparE10 strain after addition of 5 μg/ml novobiocin (Nov). Novobiocin was added at time 0, and cultures were either kept at 30°C (lanes 1–4) or shifted to 42°C (lanes 5–8). DNA was resolved on a 1% TAE gel with 10 μg/ml chloroquine. The positions of (−) supercoiled, [(−) sc], and (+) supercoiled, [(+) sc] topoisomers are shown. (B) pBR322 DNA was isolated from parC+ (Left) and parCr norfloxacin-resistant (Right) E. coli strains that had been treated with 100 μg/ml norfloxacin. The DNA was analyzed by two-dimensional agarose gel electrophoresis. The first dimension (1D) was a 1% TAE agarose gel without chloroquine, and the second dimension (2D) contained 10 μg/ml chloroquine. Reference plasmid DNA relaxed in vitro by calf thymus topoisomerase I is shown (Lower). The positions of (+) and (−) topoisomers are shown.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
