

Hypersensitivity to DNA damage leads to increased apoptosis during early mouse development
- PMID: 10950870
- PMCID: PMC316856
Hypersensitivity to DNA damage leads to increased apoptosis during early mouse development
Abstract
Gastrulation in mice is associated with the start of extreme proliferation and differentiation. The potential cost to the embryo of a very rapid proliferation rate is a high production of damaged cells. We demonstrate a novel surveillance mechanism for the elimination of cells damaged by ionizing radiation during mouse gastrulation. During this restricted developmental window, the embryo becomes hypersensitive to DNA damage induced by low dose irradiation (<0.5 Gy) and undergoes apoptosis without cell cycle arrest. Intriguingly, embryonic cells, including germ cell progenitors, but not extraembryonic cells, become hypersensitive to genotoxic stress and undergo Atm- and p53-dependent apoptosis. Thus, hypersensitivity to apoptosis in the early mouse embryo is a cell fate-dependent mechanism to ensure genomic integrity during a period of extreme proliferation and differentiation.
Figures
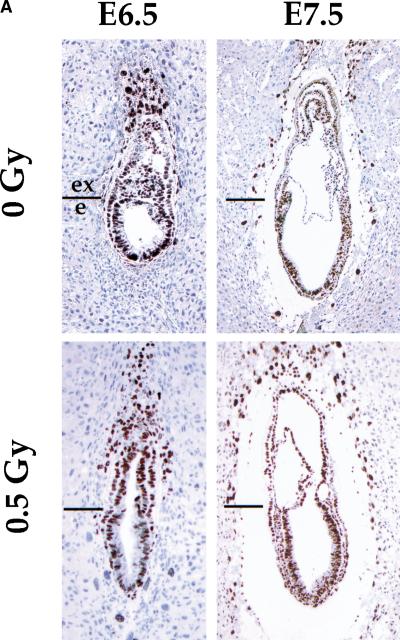

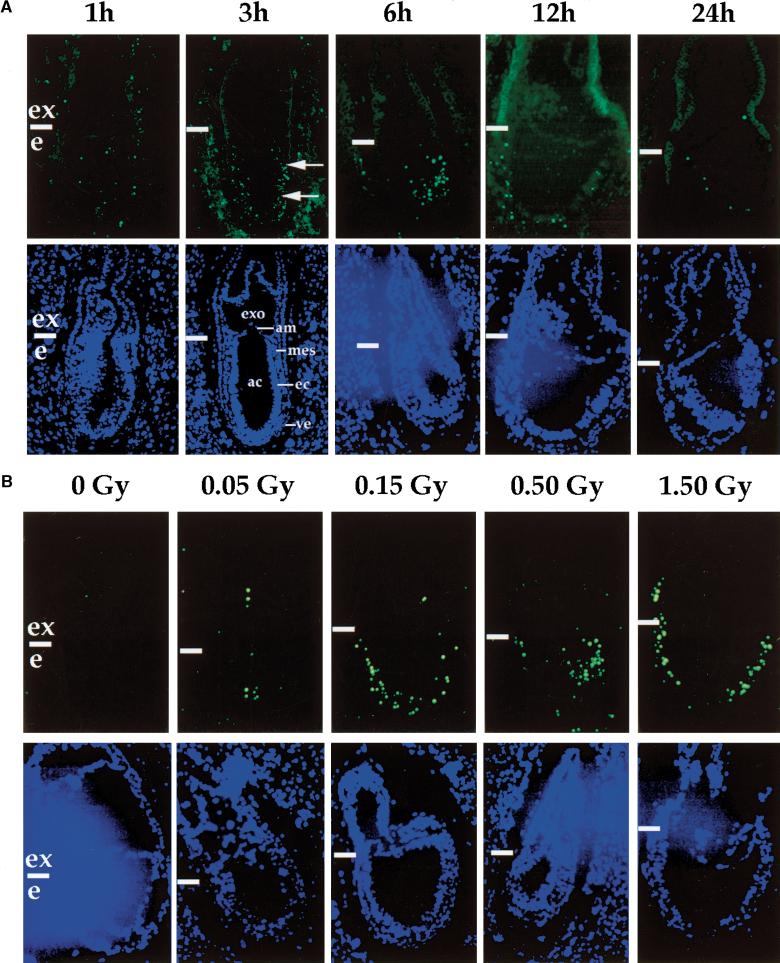
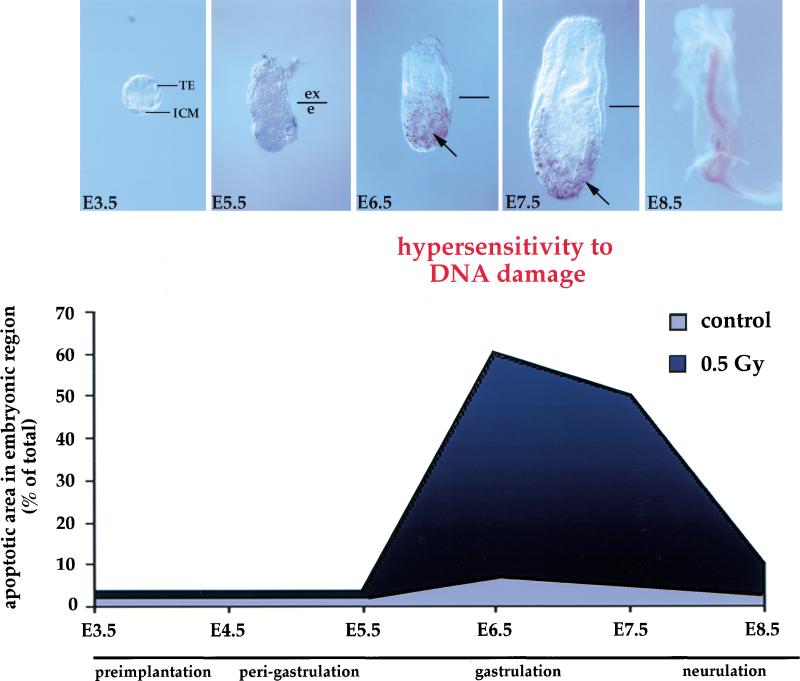

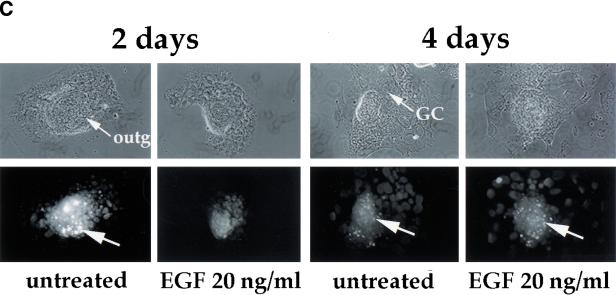
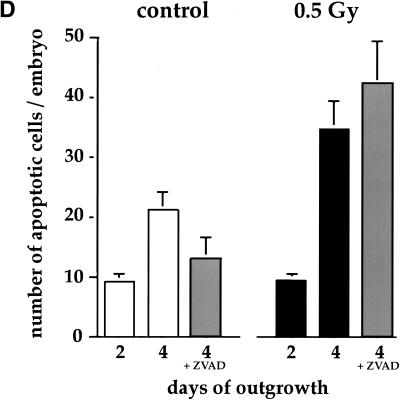
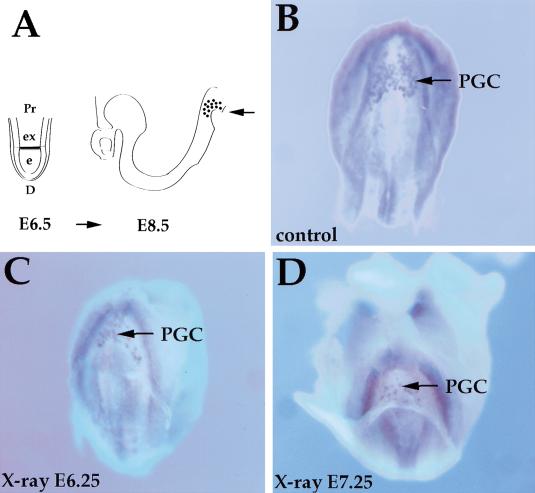
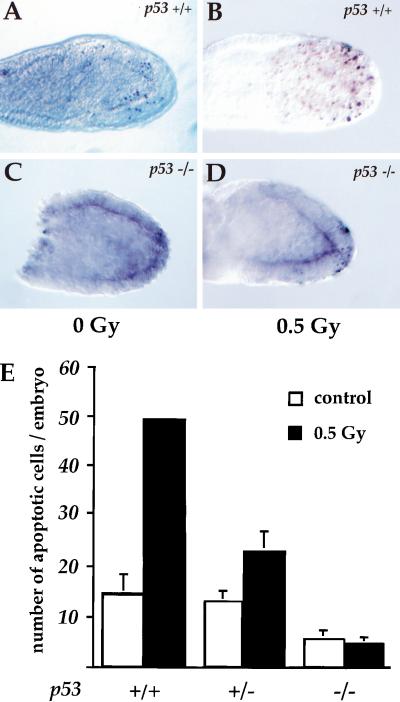


References
-
- Aladjem MI, Spike BT, Rodewald LW, Hope TJ, Klemm M, Jaenisch R, Wahl GM. ES cells do not activate p53-dependent stress responses and undergo p53-independent apoptosis in response to DNA damage. Curr Biol. 1998;8:145–155. - PubMed
-
- Armstrong JF, Kaufman MH, Harrison DJ, Clarke AR. High-frequency developmental abnormalities in p53-deficient mice. Curr Biol. 1995;5:931–936. - PubMed
-
- Banin S, Moyal L, Shieh S, Taya Y, Anderson CW, Chessa L, Smorodinsky NI, Prives C, Reiss Y, Shiloh Y, Ziv Y. Enhanced phosphorylation of p53 by ATM in response to DNA damage. Science. 1998;281:1674–1677. - PubMed
-
- Barlow C, Hirotsune S, Paylor R, Liyanage M, Eckhaus M, Collins F, Shiloh Y, Crawley JN, Ried T, Tagle D, Wynshaw-Boris A. Atm-deficient mice: A paradigm of ataxia telangiectasia. Cell. 1996;86:159–171. - PubMed
-
- Barlow C, Brown KD, Deng CX, Tagle DA, Wynshaw-Boris A. Atm selectively regulates distinct p53-dependent cell-cycle checkpoint and apoptotic pathways. Nat Genet. 1997;17:453–456. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous