

Ligand regulation of green fluorescent protein-tagged forms of the human beta(1)- and beta(2)-adrenoceptors; comparisons with the unmodified receptors
- PMID: 10952671
- PMCID: PMC1572266
- DOI: 10.1038/sj.bjp.0703506
Ligand regulation of green fluorescent protein-tagged forms of the human beta(1)- and beta(2)-adrenoceptors; comparisons with the unmodified receptors
Abstract
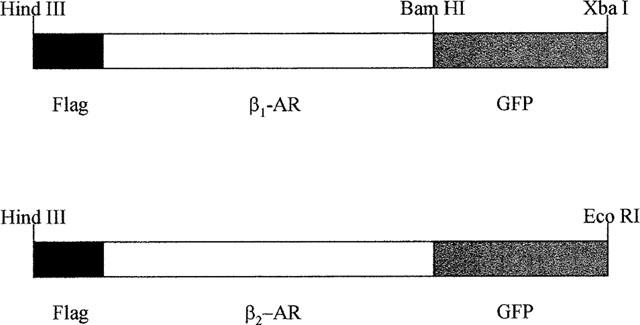
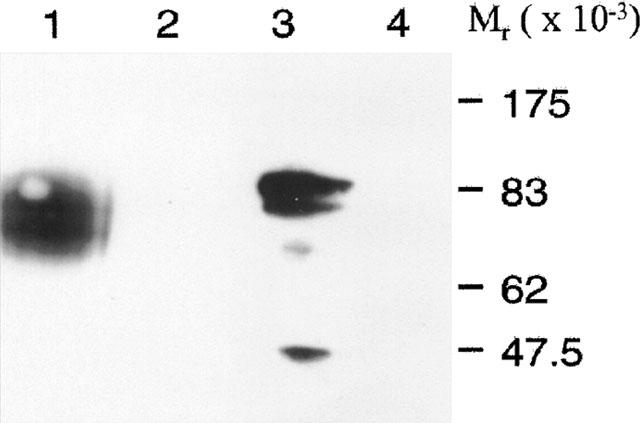
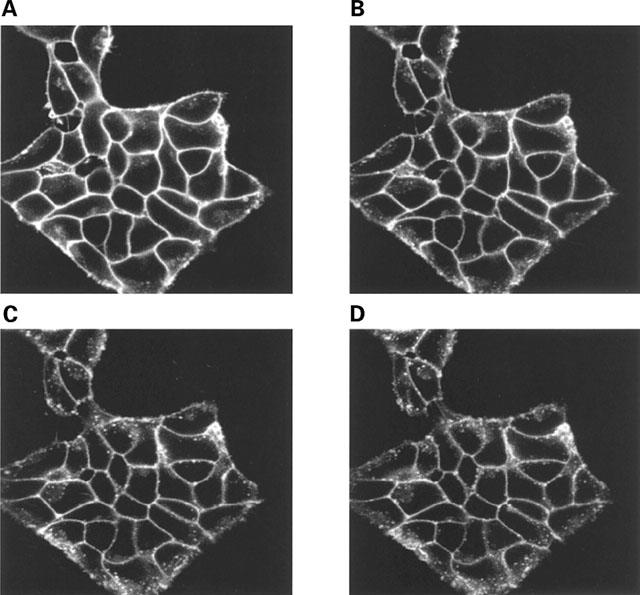
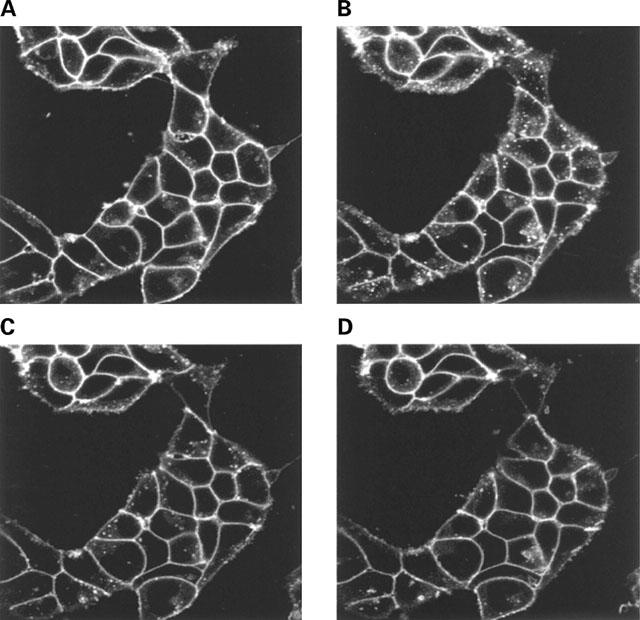
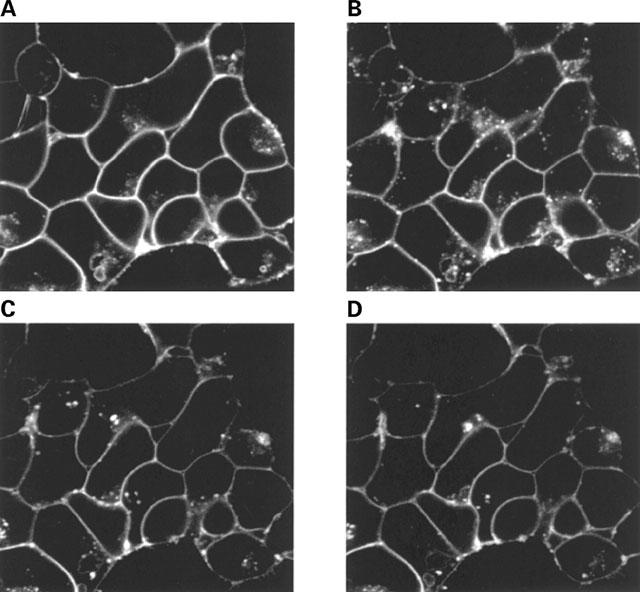
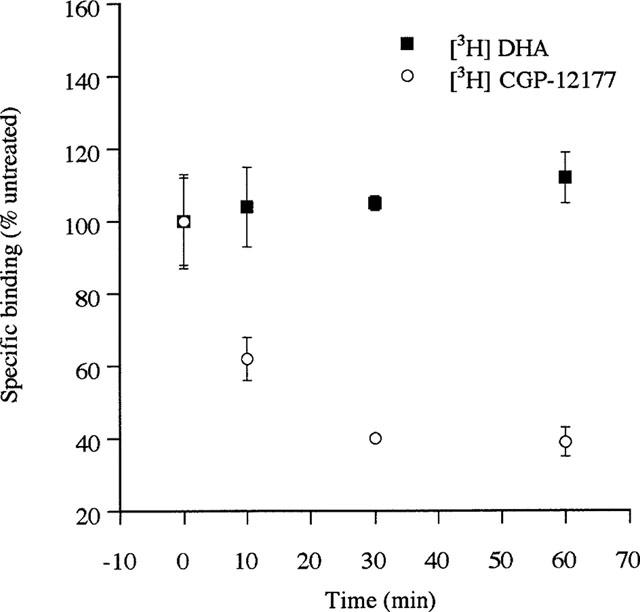
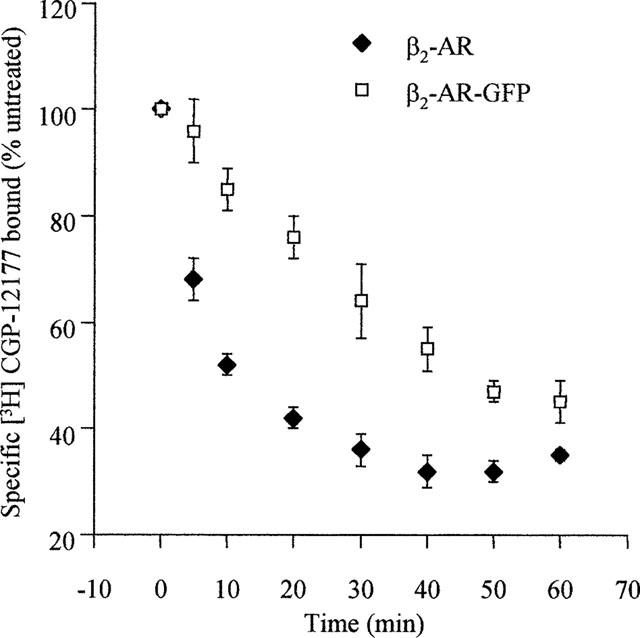
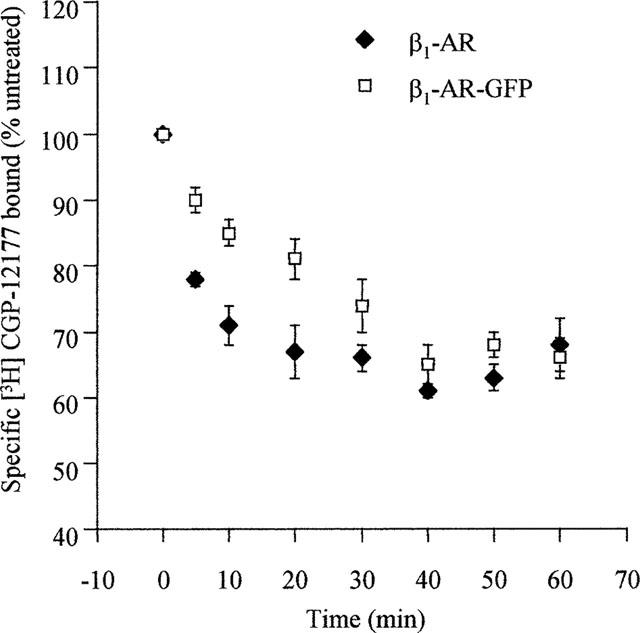
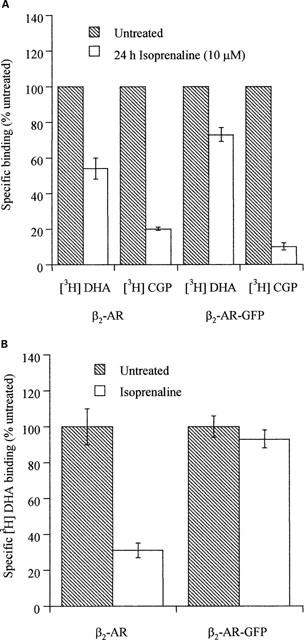
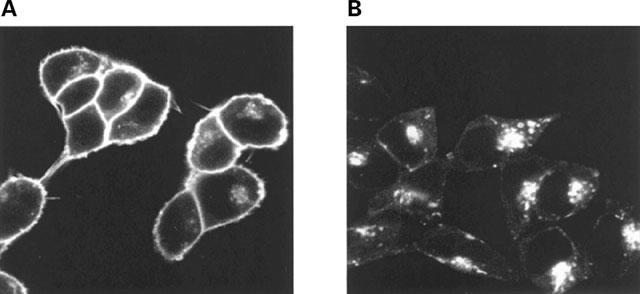
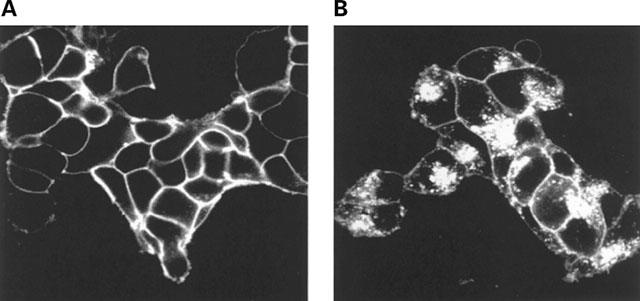
Stable clones of HEK293 cells expressing either FLAG(TM) epitope-tagged, wild type human beta(1)- and beta(2)-adrenoceptors or C-terminally green fluorescent protein (GFP)-tagged forms of these receptors were established. The binding affinity of [(3)H]-dihydroalprenolol and other ligands was little affected by addition of GFP to the C-terminal of either receptor. Isoprenaline induced the internalisation of both beta(1)-adrenoceptor-GFP and beta(2)-adrenoceptor-GFP and following removal of the agonist both constructs were able to recycle to the cell surface. The extent of internalisation of beta(2)-adrenoceptor-GFP produced by isoprenaline was substantially greater than for beta(1)-adrenoceptor-GFP. C-terminal addition of GFP slowed markedly the rate of internalization of both the beta(1)-adrenoceptor and the beta(2)-adrenoceptor in response to isoprenaline. Sustained exposure to isoprenaline (24 h) produced substantially greater levels of downregulation of native beta(2)-adrenoceptor compared to beta(2)-adrenoceptor-GFP although both were equally effectively removed from the plasma membrane. Sustained exposure to isoprenaline resulted in a large fraction of beta(2)-adrenoceptor-GFP becoming trapped in internal vesicles/lysosomes but not degraded. Even after sustained exposure to isoprenaline a significant fraction of beta(1)-adrenoceptor-GFP remained at the cell surface. These results indicate that although GFP tagging of beta-adrenoceptors can provide qualitative visual patterns of agonist-induced receptor trafficking and regulation in HEK293 cells the quantitative details vary markedly from those obtained with the unmodified receptors.
Figures
References
-
- AWAJI T., HIRASAWA H., KATAOKA M., SHINOURA H., NAKAYAMA Y., SUGAWARA T., IZUMI S., TSUJIMOTO G. Real-time optical monitoring of ligand-mediated internalisation of α1b-adrenoceptor with green fluorescent protein. Mol. Endocrinol. 1998;12:1099–1111. - PubMed
-
- BARAK L.S., FERGUSON S.S., ZHANG J., MARTENSON C., MEYER T., CARON M.G. Internal trafficking and surface mobility of a functionally intact beta2-adrenergic receptor-green fluorescent protein conjugate. Mol. Pharmacol. 1997;51:177–184. - PubMed
-
- BARLIC J., KHANDAKER M.H., MAHON E., ANDREWS J., DEVRIES M.E., MITCHELL G.B., RAHIMPOUR R., TAN C.M., FERGUSON S.S.G., KELVIN D.J. β-arrestins regulate interleukin-8-induced CXCR1 internalization. J. Biol. Chem. 1999;274:16287–16294. - PubMed
-
- CAO T.T., DEACON H.W., RECZEK D., BRETSCHER A., VON ZASTROW M. A kinase-regulated PDZ-domain interaction controls endocytic sorting of the β2-adrenergic receptor. Nature. 1999;401:286–290. - PubMed
-
- CONWAY B.R., MINOR L.K., XU J.Z., GUNNET J.W., DEBIASIO R., D'ANDREA M.R., RUBIN R., DEBIASIO R., GIULIANO K., ZHOU L., DEMAREST K.T. Quantification of G-protein coupled receptor internalization using G-protein coupled receptor-green fluorescent protein conjugates with the Array ScanTM high-content screening system. J. Biomol. Screening. 1999;4:75–86. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
