

Leptin enhances wound re-epithelialization and constitutes a direct function of leptin in skin repair
- PMID: 10953025
- PMCID: PMC380250
- DOI: 10.1172/JCI9148
Leptin enhances wound re-epithelialization and constitutes a direct function of leptin in skin repair
Abstract
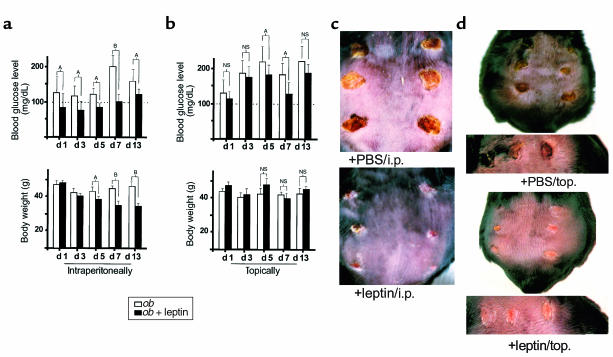
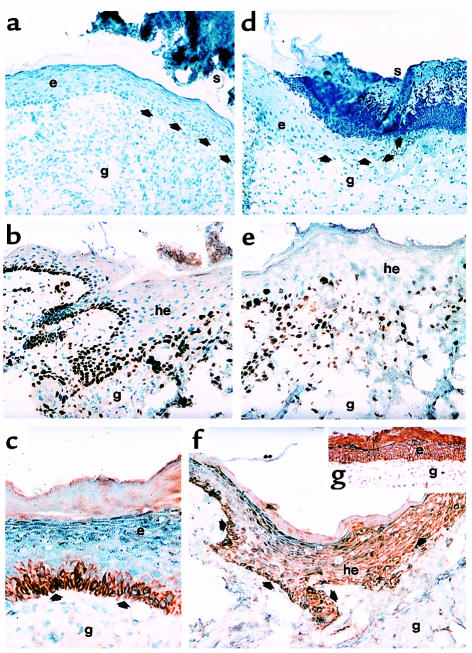
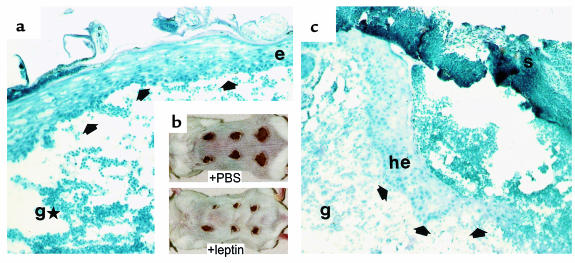
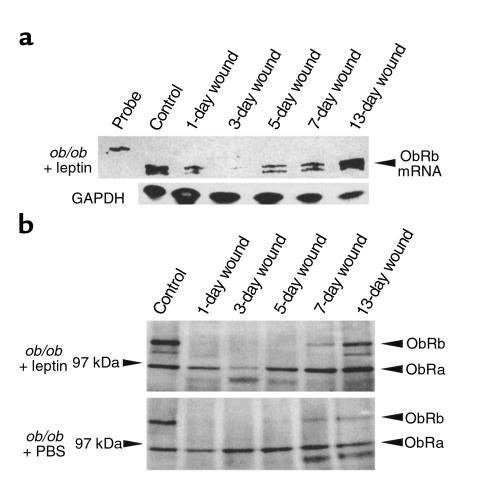
Wound-healing disorders are a therapeutic problem of extensive clinical importance. Leptin-deficient ob/ob mice are characterized by a severely delayed wound healing that has been explained by the mild diabetic phenotype of these animals. Here we demonstrate that systemically and topically supplemented leptin improved re-epithelialization of wounds in ob/ob mice. Leptin completely reversed the atrophied morphology of the migrating epithelial tongue observed at the wound margins of leptin-deficient animals into a well-organized hyperproliferative epithelium. Moreover, topically supplemented leptin accelerated normal wound-healing conditions in wild-type mice. As assessed by immunohistochemistry, proliferating keratinocytes located at the wound margins specifically expressed the leptin-receptor subtype ObRb during repair. Additionally, leptin mediated a mitogenic stimulus to the human keratinocyte cell line HaCaT and human primary keratinocytes in vitro. Therefore, leptin might represent an effective novel therapeutic factor to improve impaired wound-healing conditions.
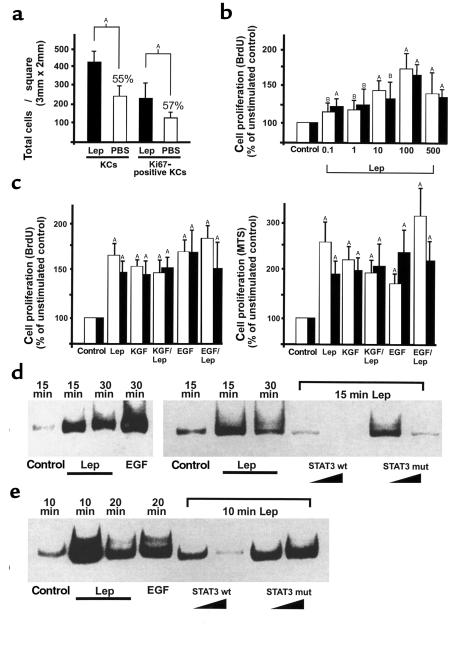
Figures
References
-
- Zhang Y, et al. Positional cloning of the mouse obese gene and its human homologue. Nature. 1994;372:425–432. - PubMed
-
- Masuzaki H, et al. Human obese gene expression. Adipocyte-specific expression and regional differences in the adipose tissue. Diabetes. 1995;44:855–858. - PubMed
-
- Halaas JL, et al. Weight-reducing effects of the plasma protein encoded by the obese gene. Science. 1995;269:543–546. - PubMed
-
- Vaisse C, et al. Leptin activation of STAT3 in the hypothalamus of wild-type and ob/ob mice but not db/db mice. Nat Genet. 1996;14:95–97. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
