

Mutants of human immunodeficiency virus type 1 (HIV-1) reverse transcriptase resistant to nonnucleoside reverse transcriptase inhibitors demonstrate altered rates of RNase H cleavage that correlate with HIV-1 replication fitness in cell culture
- PMID: 10954539
- PMCID: PMC116350
- DOI: 10.1128/jvi.74.18.8390-8401.2000
Mutants of human immunodeficiency virus type 1 (HIV-1) reverse transcriptase resistant to nonnucleoside reverse transcriptase inhibitors demonstrate altered rates of RNase H cleavage that correlate with HIV-1 replication fitness in cell culture
Abstract
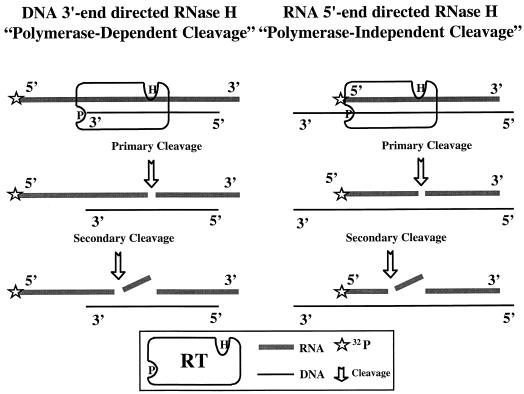
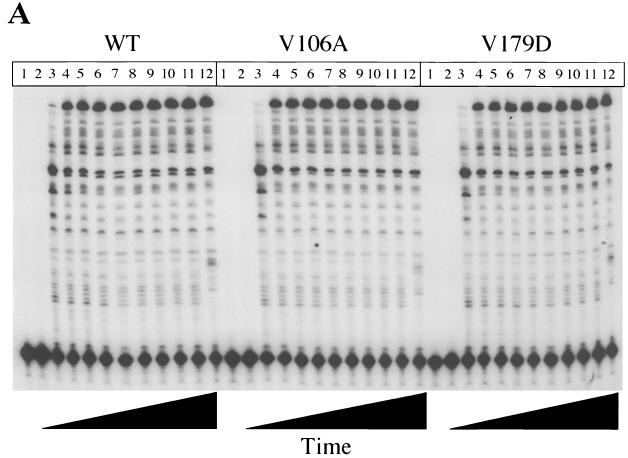
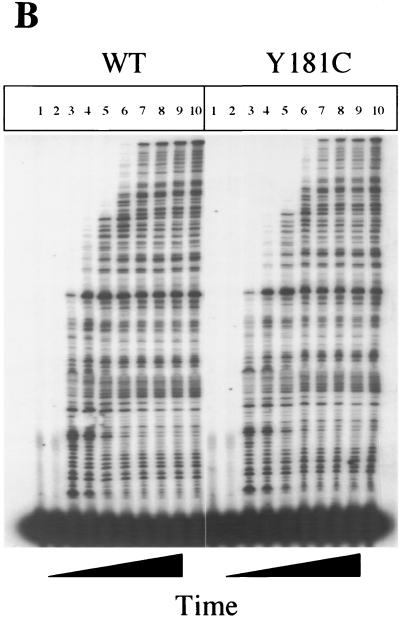
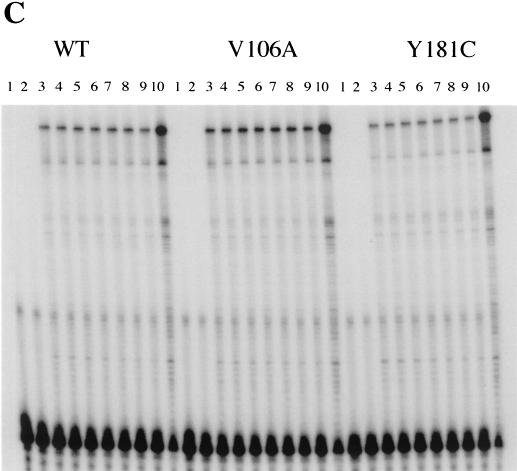
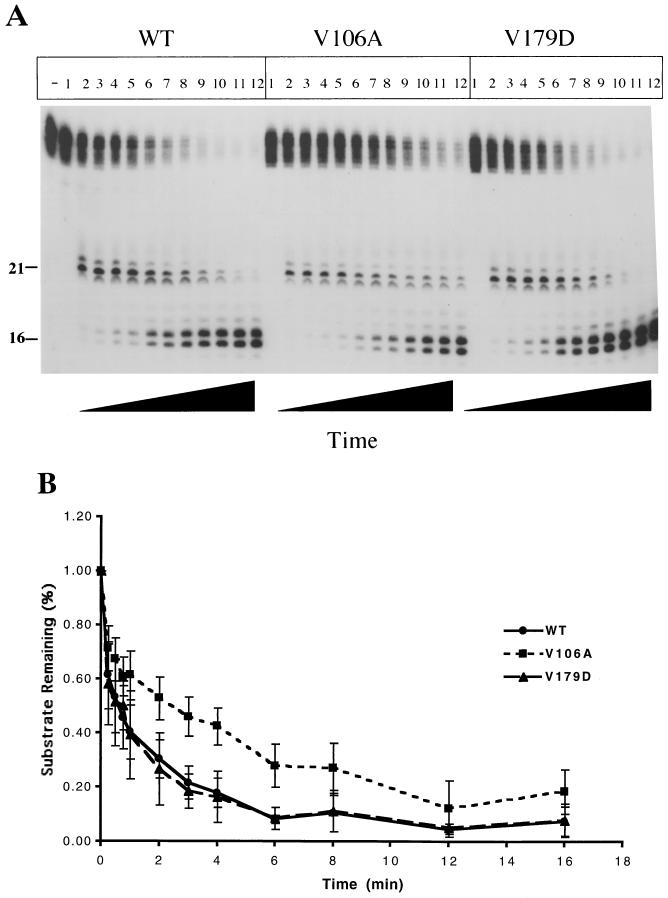
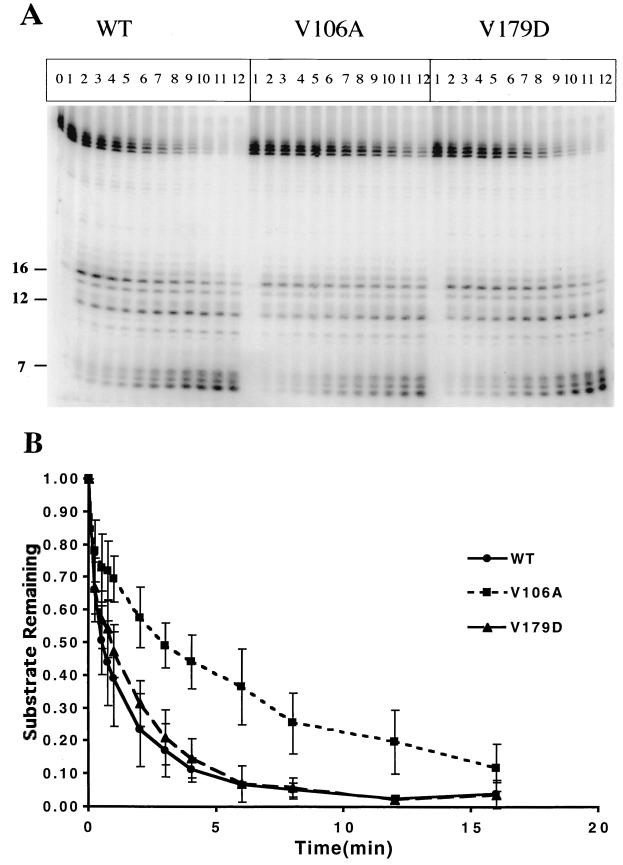
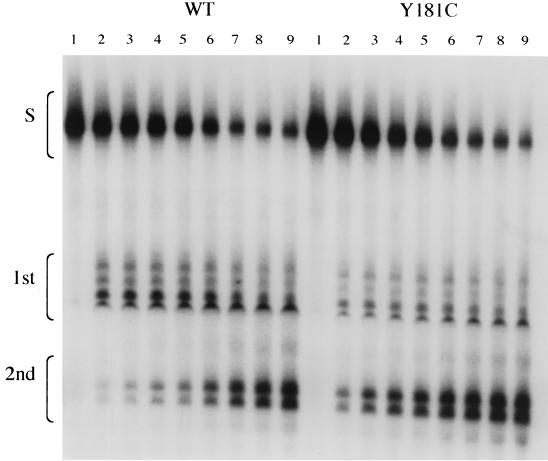
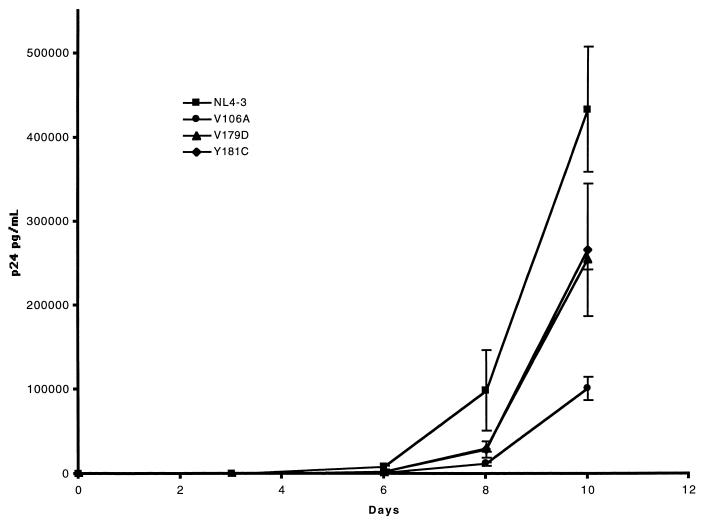
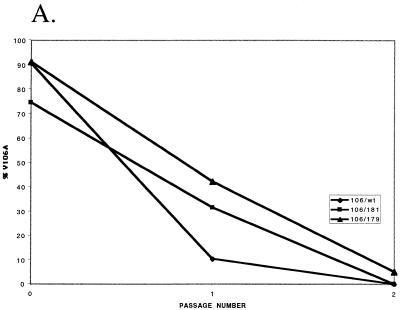
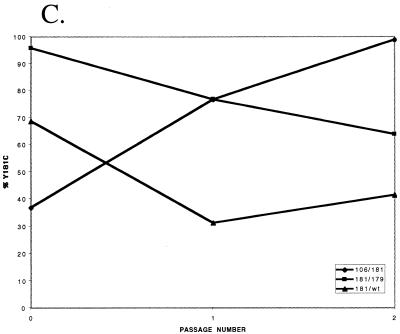
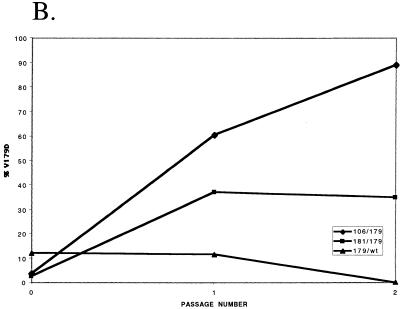
Three mutants of human immunodeficiency virus type 1 (HIV-1) reverse transcriptase (V106A, V179D, and Y181C), which occur in clinical isolates and confer resistance to nonnucleoside reverse transcriptase inhibitors (NNRTIs), were analyzed for RNA- and DNA-dependent DNA polymerization and RNase H cleavage. All mutants demonstrated processivities of polymerization that were indistinguishable from wild-type enzyme under conditions in which deoxynucleoside triphosphates were not limiting. The V106A reverse transcriptase demonstrated a three- to fourfold slowing of both DNA 3'-end-directed and RNA 5'-end-directed RNase H cleavage relative to both wild-type and V179D enzymes, similar to what was observed for P236L in a previously published study (P. Gerondelis et al., J. Virol. 73:5803-5813, 1999). In contrast, the Y181C reverse transcriptase demonstrated a selective acceleration of the secondary RNase H cleavage step during both modes of RNase H cleavage. The relative replication fitness of these mutants in H9 cells was assessed in parallel infections as well as in growth competition experiments. Of the NNRTI-resistant mutants, V179D was more fit than Y181C, and both of these mutants were more fit than V106A, which demonstrated the greatest reduction in RNase H cleavage. These findings, in combination with results from previous work, suggest that abnormalities in RNase H cleavage are a common characteristic of HIV-1 mutants resistant to NNRTIs and that combined reductions in the rates of DNA 3'-end- and RNA 5'-end-directed cleavages are associated with significant reductions in the replication fitness of HIV-1.
Figures
References
-
- Amacker M, Hubscher U. Chimeric HIV-1 and feline immunodeficiency virus reverse transcriptases: critical role of the p51 subunit in the structural integrity of heterodimeric lentiviral DNA polymerases. J Mol Biol. 1998;278:757–765. - PubMed
-
- Arion D, Kaushik N, McCormick S, Borkow G, Parniak M A. Phenotypic mechanism of HIV-1 resistance to 3′-azido-3′-deoxythymidine (AZT): increased polymerization processivity and enhanced sensitivity to pyrophosphate of the mutant viral reverse transcriptase. Biochemistry. 1998;37:15908–15917. - PubMed
-
- Arts E J, Le Grice S F. Interaction of retroviral reverse transcriptase with template-primer duplexes during replication. Prog Nucleic Acid Res Mol Biol. 1998;58:339–393. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
