

Characterization of the role of AMP-activated protein kinase in the regulation of glucose-activated gene expression using constitutively active and dominant negative forms of the kinase
- PMID: 10958668
- PMCID: PMC86183
- DOI: 10.1128/MCB.20.18.6704-6711.2000
Characterization of the role of AMP-activated protein kinase in the regulation of glucose-activated gene expression using constitutively active and dominant negative forms of the kinase
Abstract
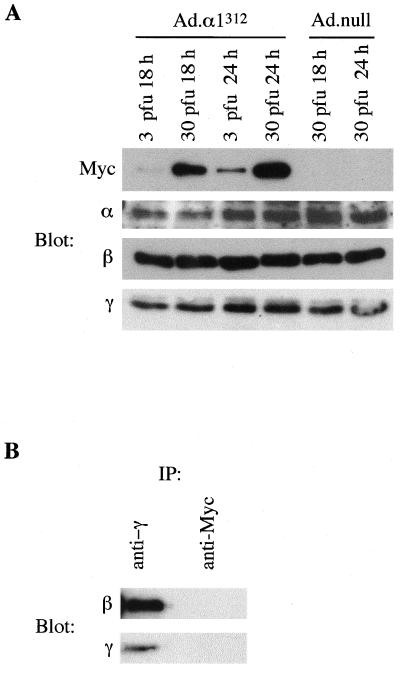
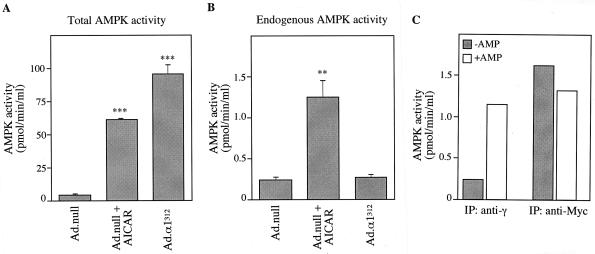
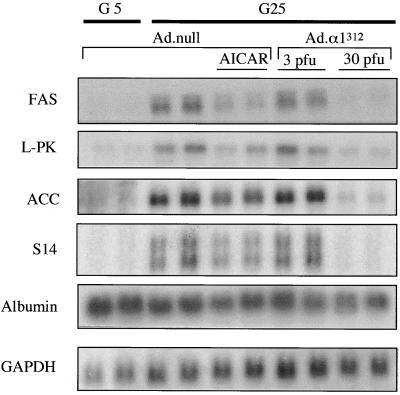
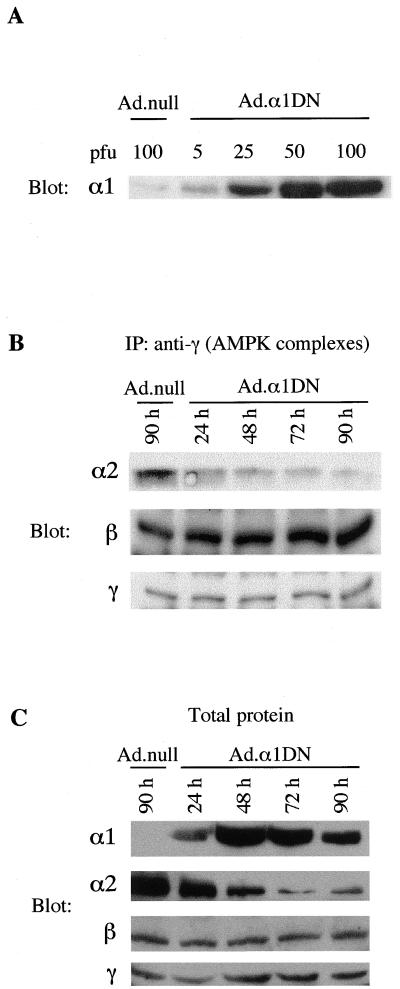
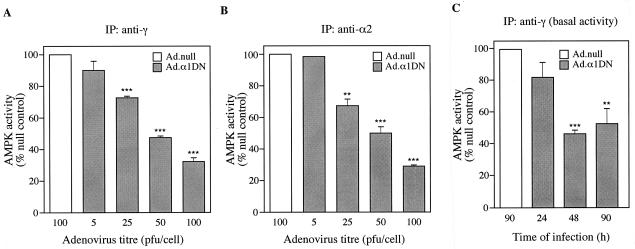
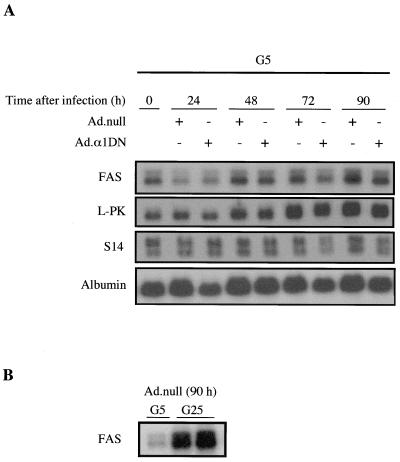
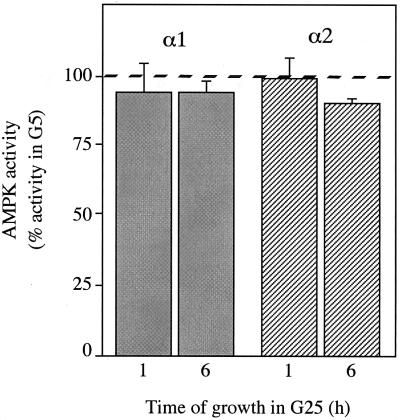
In the liver, glucose induces the expression of a number of genes involved in glucose and lipid metabolism, e.g., those encoding L-type pyruvate kinase and fatty acid synthase. Recent evidence has indicated a role for the AMP-activated protein kinase (AMPK) in the inhibition of glucose-activated gene expression in hepatocytes. It remains unclear, however, whether AMPK is involved in the glucose induction of these genes. In order to study further the role of AMPK in regulating gene expression, we have generated two mutant forms of AMPK. One of these (alpha1(312)) acts as a constitutively active kinase, while the other (alpha1DN) acts as a dominant negative inhibitor of endogenous AMPK. We have used adenovirus-mediated gene transfer to express these mutants in primary rat hepatocytes in culture in order to determine their effect on AMPK activity and the transcription of glucose-activated genes. Expression of alpha1(312) increased AMPK activity in hepatocytes and blocked completely the induction of a number of glucose-activated genes in response to 25 mM glucose. This effect is similar to that observed following activation of AMPK by 5-amino-imidazolecarboxamide riboside. Expression of alpha1DN markedly inhibited both basal and stimulated activity of endogenous AMPK but had no effect on the transcription of glucose-activated genes. Our results suggest that AMPK is involved in the inhibition of glucose-activated gene expression but not in the induction pathway. This study demonstrates that the two mutants we have described will provide valuable tools for studying the wider physiological role of AMPK.
Figures
References
-
- Carling D, Aguan K, Woods A, Verhoeven A J M, Beri R K, Brennan C H, Sidebottom C, Davison M D, Scott J. Mammalian AMP-activated protein kinase is homologous to yeast and plant protein kinases involved in the regulation of carbon metabolism. J Biol Chem. 1994;269:11442–11448. - PubMed
-
- Celenza J L, Carlson M. A yeast gene that is essential for release from glucose repression encodes a protein kinase. Science. 1986;233:1175–1180. - PubMed
-
- Chomczynski P, Sacchi N. Single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction. Anal Biochem. 1987;162:156–159. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources