

The N terminus of the centromere H3-like protein Cse4p performs an essential function distinct from that of the histone fold domain
- PMID: 10958698
- PMCID: PMC88778
- DOI: 10.1128/MCB.20.18.7037-7048.2000
The N terminus of the centromere H3-like protein Cse4p performs an essential function distinct from that of the histone fold domain
Abstract
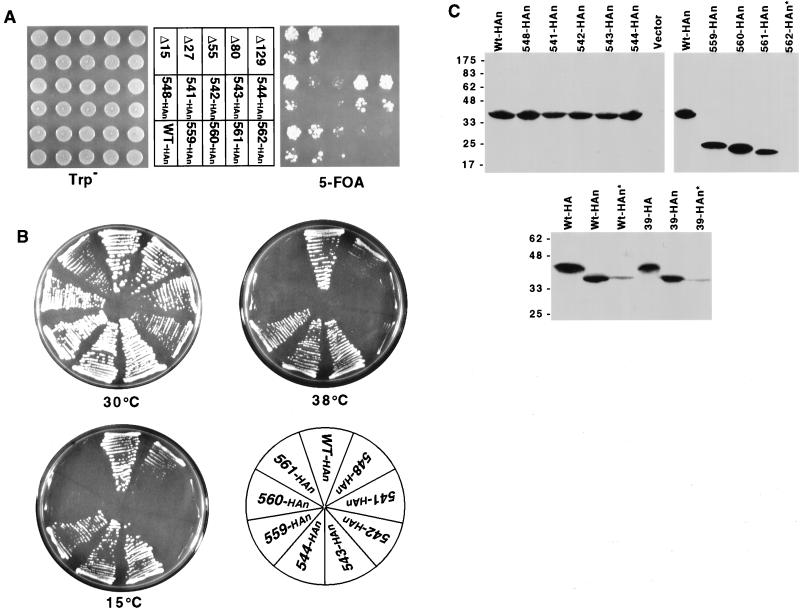
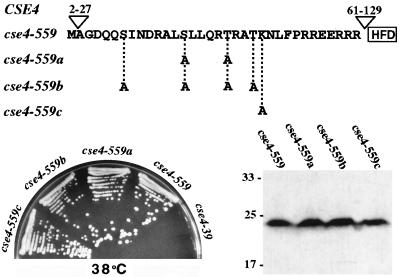
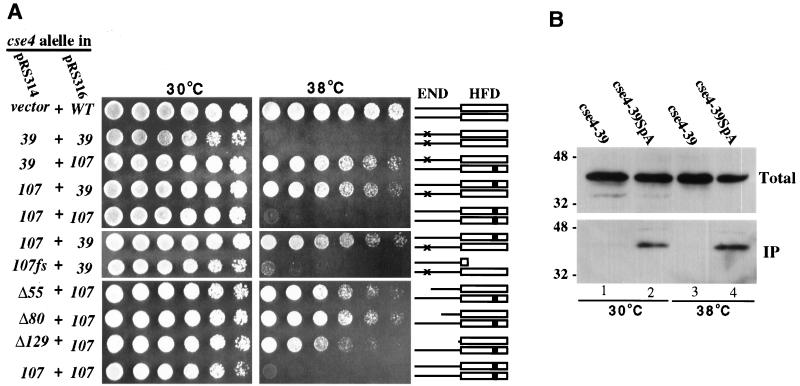
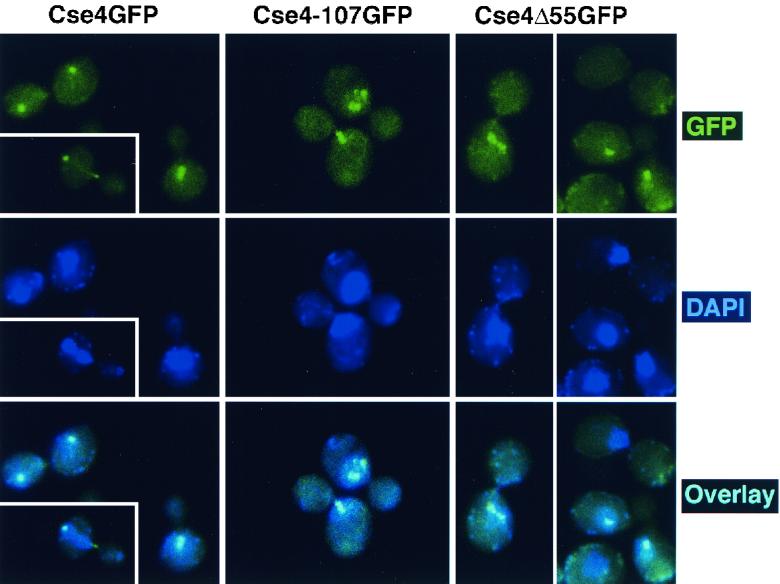
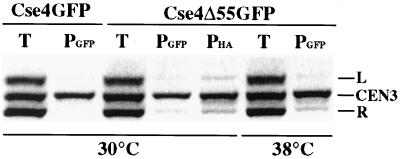
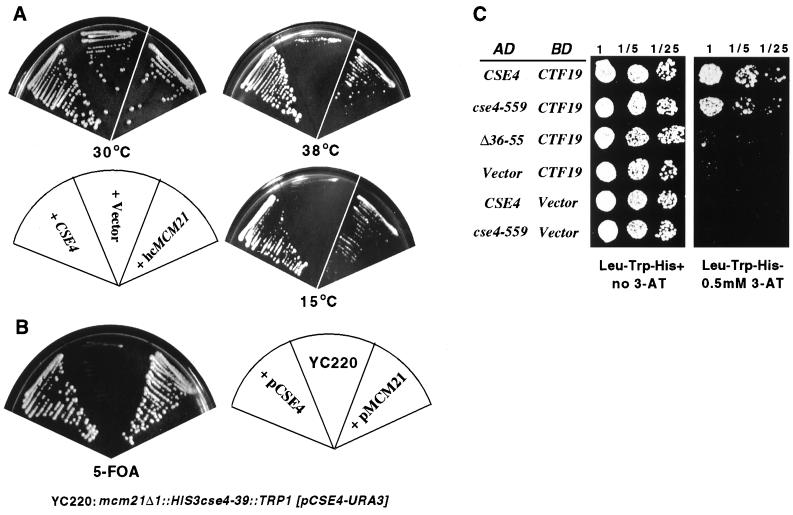
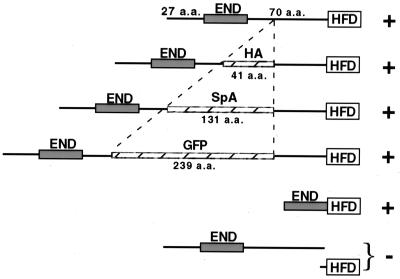
Cse4p is an evolutionarily conserved histone H3-like protein that is thought to replace H3 in a specialized nucleosome at the yeast (Saccharomyces cerevisiae) centromere. All known yeast, worm, fly, and human centromere H3-like proteins have highly conserved C-terminal histone fold domains (HFD) but very different N termini. We have carried out a comprehensive and systematic mutagenesis of the Cse4p N terminus to analyze its function. Surprisingly, only a 33-amino-acid domain within the 130-amino-acid-long N terminus is required for Cse4p N-terminal function. The spacing of the essential N-terminal domain (END) relative to the HFD can be changed significantly without an apparent effect on Cse4p function. The END appears to be important for interactions between Cse4p and known kinetochore components, including the Ctf19p/Mcm21p/Okp1p complex. Genetic and biochemical evidence shows that Cse4p proteins interact with each other in vivo and that nonfunctional cse4 END and HFD mutant proteins can form functional mixed complexes. These results support different roles for the Cse4p N terminus and the HFD in centromere function and are consistent with the proposed Cse4p nucleosome model. The structure-function characteristics of the Cse4p N terminus are relevant to understanding how other H3-like proteins, such as the human homolog CENP-A, function in kinetochore assembly and chromosome segregation.
Figures

References
-
- Baker R E, Fitzgerald-Hayes M, O'Brien T C. Purification of the yeast centromere binding protein CP1 and a mutational analysis of its binding site. J Biol Chem. 1989;264:10843–10850. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases