

Lesions in gshA (Encoding gamma-L-glutamyl-L-cysteine synthetase) prevent aerobic synthesis of thiamine in Salmonella enterica serovar typhimurium LT2
- PMID: 10960103
- PMCID: PMC94667
- DOI: 10.1128/JB.182.18.5180-5187.2000
Lesions in gshA (Encoding gamma-L-glutamyl-L-cysteine synthetase) prevent aerobic synthesis of thiamine in Salmonella enterica serovar typhimurium LT2
Abstract
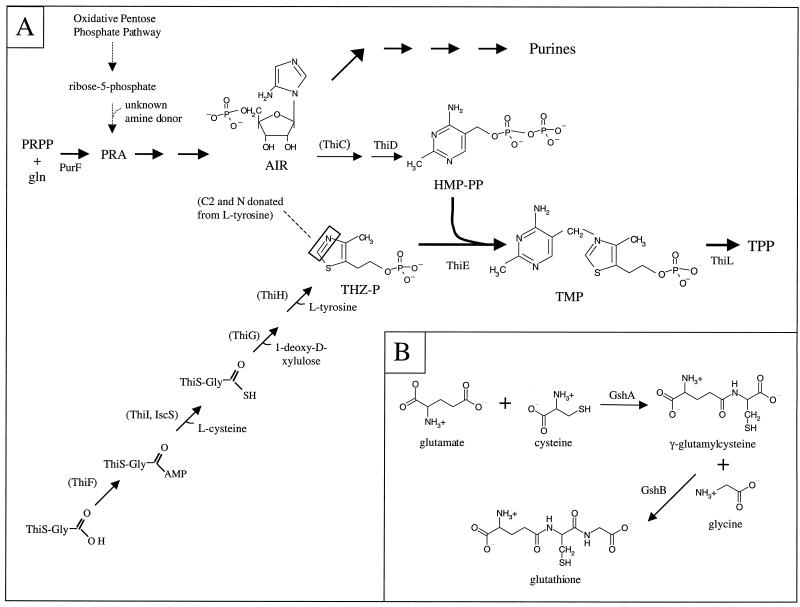
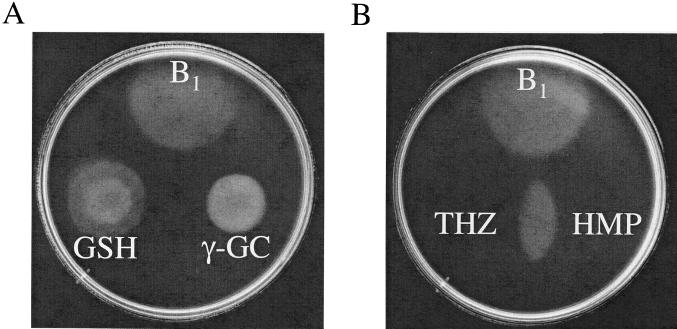
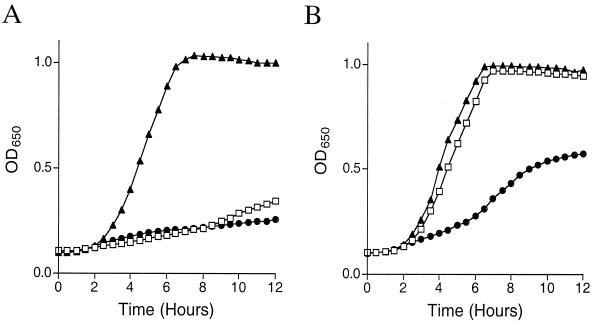
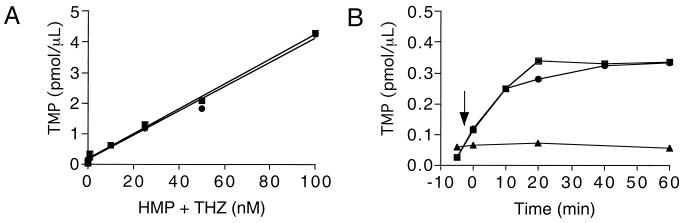
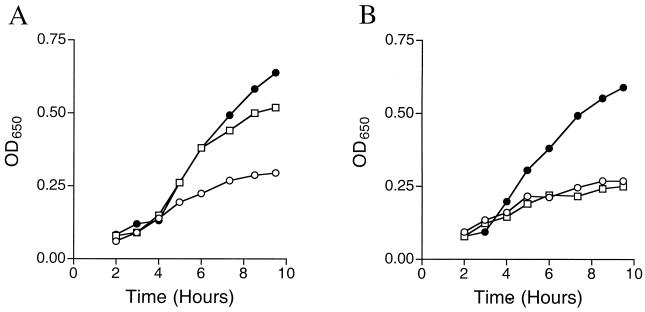
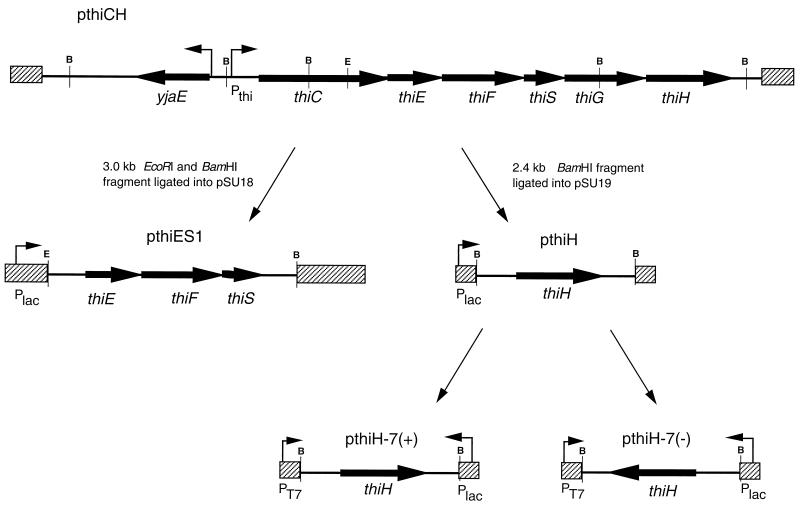
Thiamine pyrophosphate is an essential cofactor that is synthesized de novo in Salmonella enterica serovar Typhimurium and other bacteria. In addition to genes encoding enzymes in the biosynthetic pathway, mutations in other metabolic loci have been shown to prevent thiamine synthesis. The latter loci identify the integration of the thiamine biosynthetic pathway with other metabolic processes and can be uncovered when thiamine biosynthesis is challenged. Mutations in gshA, encoding gamma-L-glutamyl-L-cysteine synthetase, prevent the synthesis of glutathione, the major free thiol in the cell, and are shown here to result in a thiamine auxotrophy in some of the strains tested, including S. enterica LT2. Phenotypic characterization of the gshA mutants indicated they were similar enough to apbC and apbE mutants to warrant the definition of a class of mutants unified by (i) a requirement for both the hydroxymethyl pyrimidine (HMP) and thiazole (THZ) moiety of thiamine, (ii) the ability of L-tryosine to satisfy the THZ requirement, (iii) suppression of the thiamine requirement by anaerobic growth, and (iv) suppression by a second-site mutation at a single locus. Genetic data indicated that a defective ThiH generates the THZ requirement in these strains, and we suggest this defect is due to a reduced ability to repair a critical [Fe-S] cluster.
Figures

Similar articles
-
Mutations in apbC (mrp) prevent function of the alternative pyrimidine biosynthetic pathway in Salmonella typhimurium.J Bacteriol. 1996 Oct;178(19):5676-82. doi: 10.1128/jb.178.19.5676-5682.1996. J Bacteriol. 1996. PMID: 8824612 Free PMC article.
-
The apbE gene encodes a lipoprotein involved in thiamine synthesis in Salmonella typhimurium.J Bacteriol. 1998 Feb;180(4):885-91. doi: 10.1128/JB.180.4.885-891.1998. J Bacteriol. 1998. PMID: 9473043 Free PMC article.
-
Glutathione is required for maximal transcription of the cobalamin biosynthetic and 1,2-propanediol utilization (cob/pdu) regulon and for the catabolism of ethanolamine, 1,2-propanediol, and propionate in Salmonella typhimurium LT2.J Bacteriol. 1995 Oct;177(19):5434-9. doi: 10.1128/jb.177.19.5434-5439.1995. J Bacteriol. 1995. PMID: 7559326 Free PMC article.
-
Metabolic flux in both the purine mononucleotide and histidine biosynthetic pathways can influence synthesis of the hydroxymethyl pyrimidine moiety of thiamine in Salmonella enterica.J Bacteriol. 2002 Nov;184(22):6130-7. doi: 10.1128/JB.184.22.6130-6137.2002. J Bacteriol. 2002. PMID: 12399482 Free PMC article.
-
Mutational analysis of ThiH, a member of the radical S-adenosylmethionine (AdoMet) protein superfamily.J Biol Chem. 2004 Sep 24;279(39):40505-10. doi: 10.1074/jbc.M403985200. Epub 2004 Jul 22. J Biol Chem. 2004. PMID: 15271986
Cited by
-
Lack of YggX results in chronic oxidative stress and uncovers subtle defects in Fe-S cluster metabolism in Salmonella enterica.J Bacteriol. 2004 Nov;186(22):7626-34. doi: 10.1128/JB.186.22.7626-7634.2004. J Bacteriol. 2004. PMID: 15516576 Free PMC article.
-
Thiamine biosynthesis can be used to dissect metabolic integration.Trends Microbiol. 2010 Jun;18(6):240-7. doi: 10.1016/j.tim.2010.03.003. Epub 2010 Apr 8. Trends Microbiol. 2010. PMID: 20382023 Free PMC article.
-
Altered pathway routing in a class of Salmonella enterica serovar Typhimurium mutants defective in aminoimidazole ribonucleotide synthetase.J Bacteriol. 2001 Apr;183(7):2234-40. doi: 10.1128/JB.183.7.2234-2240.2001. J Bacteriol. 2001. PMID: 11244062 Free PMC article.
-
Radical S-adenosylmethionine enzymes.Chem Rev. 2014 Apr 23;114(8):4229-317. doi: 10.1021/cr4004709. Epub 2014 Jan 29. Chem Rev. 2014. PMID: 24476342 Free PMC article. Review. No abstract available.
-
Analysis of yggX and gshA mutants provides insights into the labile iron pool in Salmonella enterica.J Bacteriol. 2008 Dec;190(23):7608-13. doi: 10.1128/JB.00639-08. Epub 2008 Oct 3. J Bacteriol. 2008. PMID: 18835989 Free PMC article.
References
-
- Altschul S F, Gish W, Miller W, Myers E W, Lipman D J. Basic local alignment search tool. J Mol Biol. 1990;215:403–410. - PubMed
-
- Apontoweil P, Berends W. Mapping of gshA, a gene for the biosynthesis of glutathione in Escherichia coli K12. Mol Gen Genet. 1975;141:91–95. - PubMed
-
- Begley T P, Downs D M, Ealick S, McLafferty F, van Loon D, Taylor S, Chiu H, Kinsland C, Reddick J, Xi J, Campobasso N. Thiamin synthesis in prokaryotes. Arch Microbiol. 1999;171:293–300. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
