

Coding of border ownership in monkey visual cortex
- PMID: 10964965
- PMCID: PMC4784717
- DOI: 10.1523/JNEUROSCI.20-17-06594.2000
Coding of border ownership in monkey visual cortex
Abstract
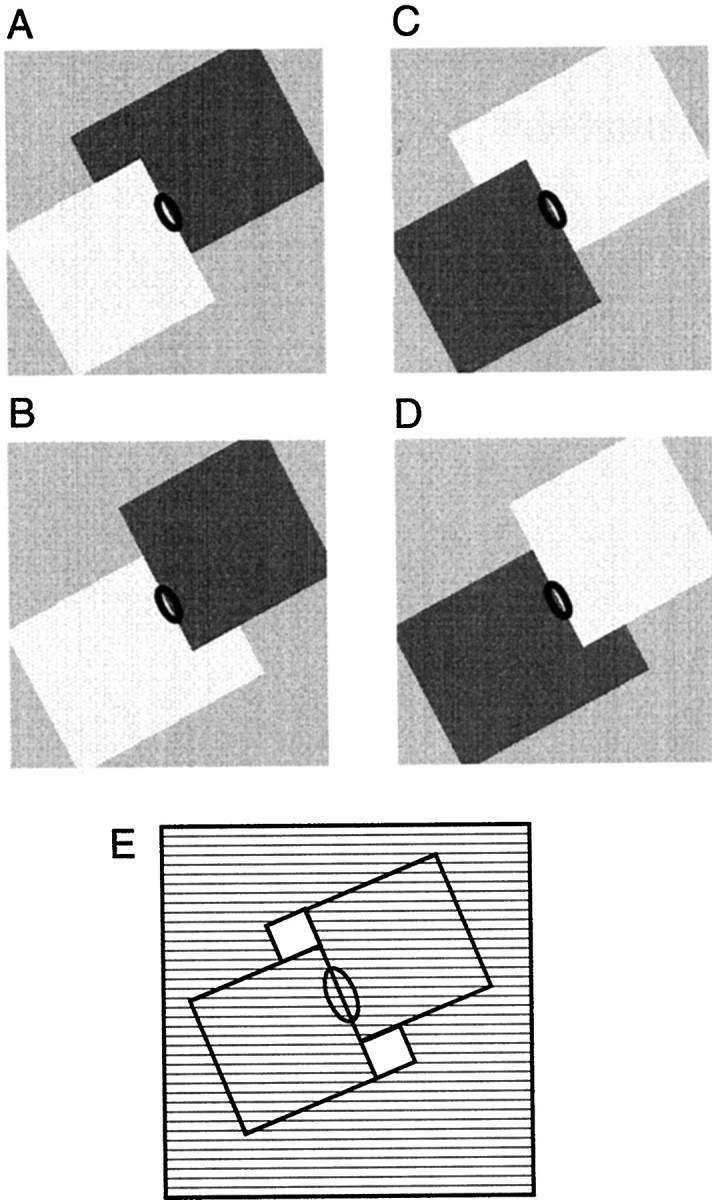
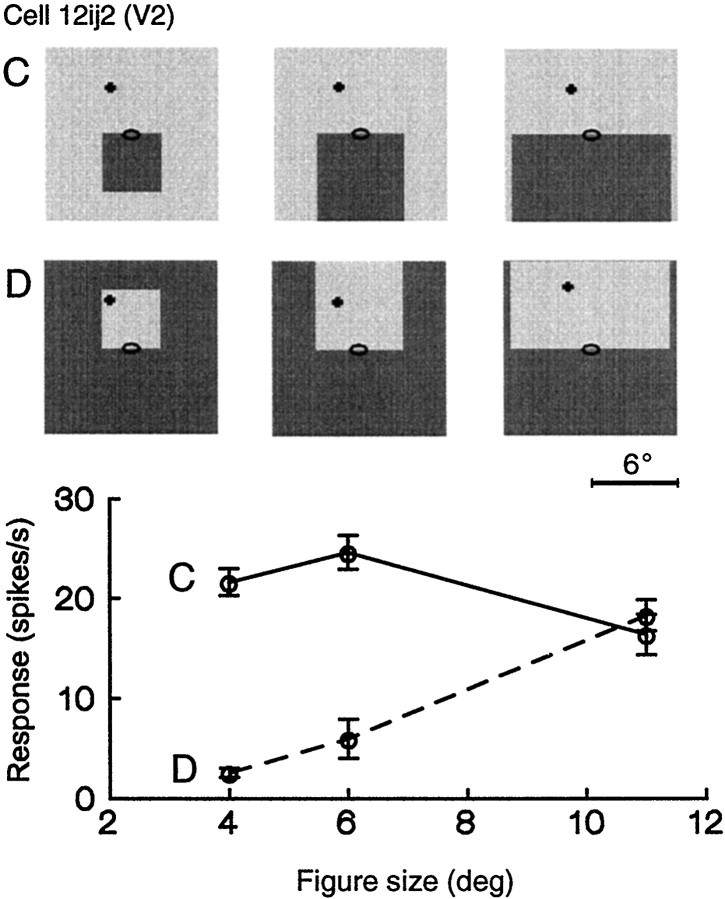
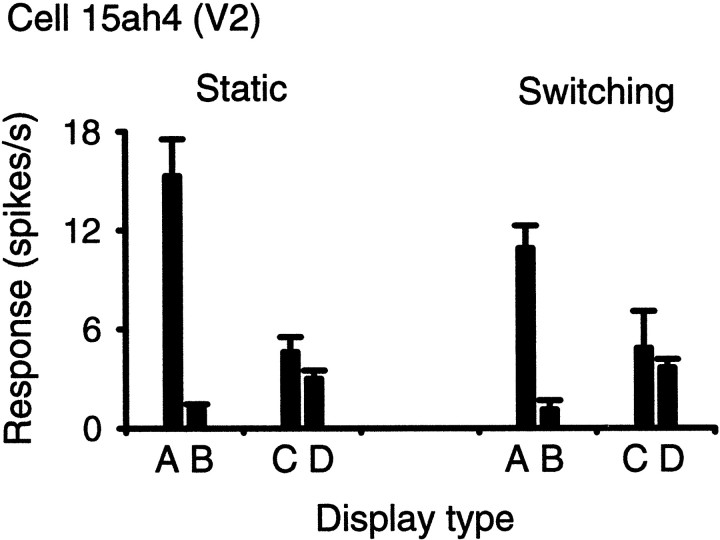
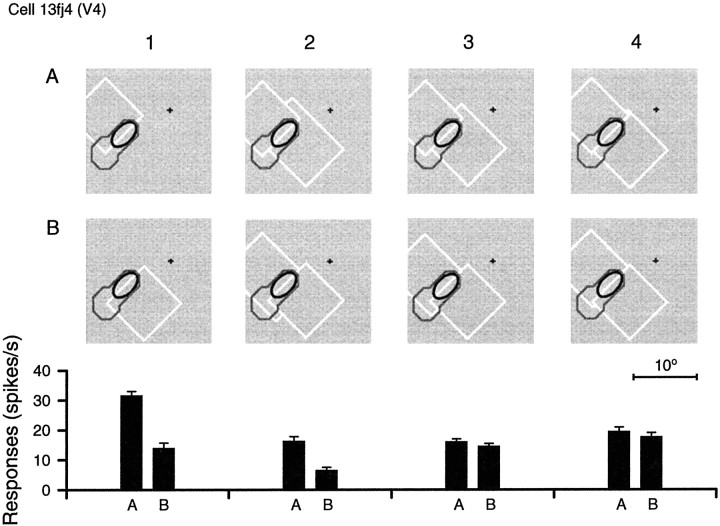
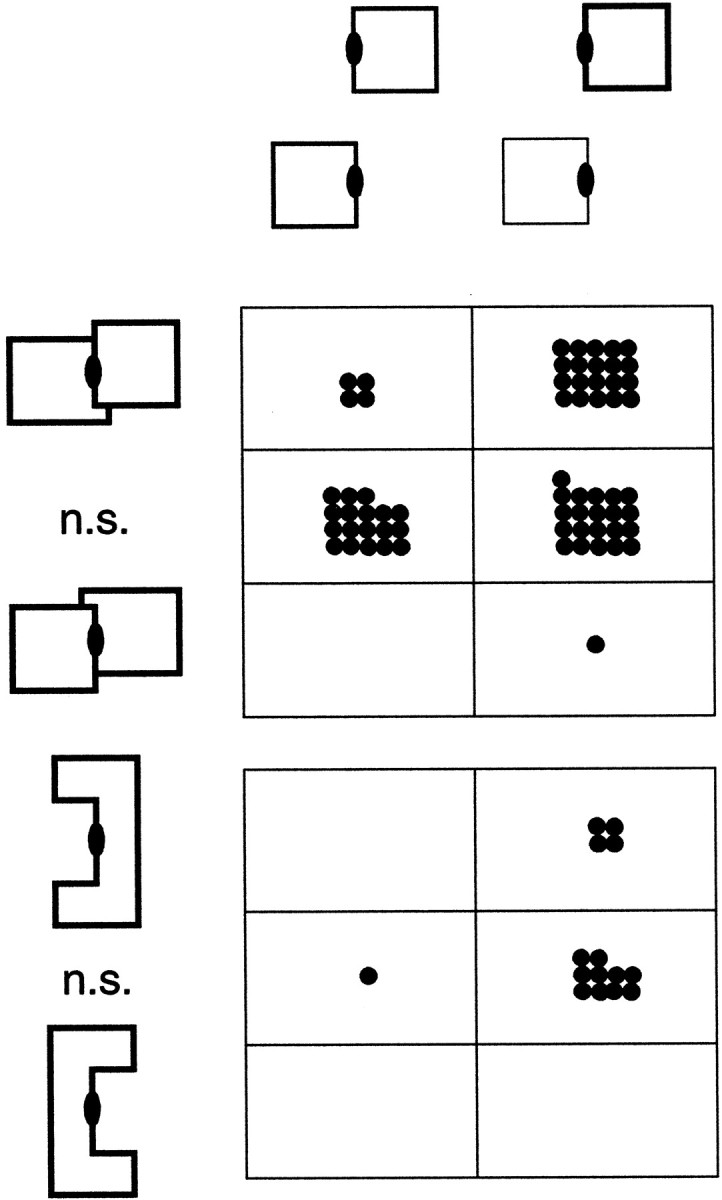
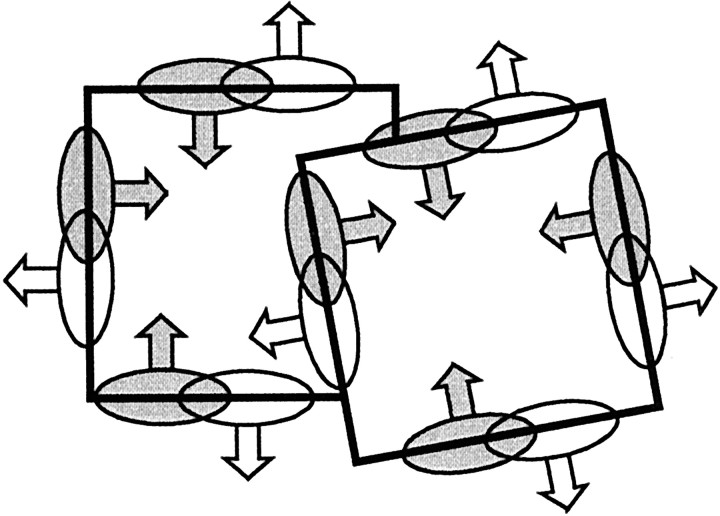
Areas V1 and V2 of the visual cortex have traditionally been conceived as stages of local feature representations. We investigated whether neural responses carry information about how local features belong to objects. Single-cell activity was recorded in areas V1, V2, and V4 of awake behaving monkeys. Displays were used in which the same local feature (contrast edge or line) could be presented as part of different figures. For example, the same light-dark edge could be the left side of a dark square or the right side of a light square. Each display was also presented with reversed contrast. We found significant modulation of responses as a function of the side of the figure in >50% of neurons of V2 and V4 and in 18% of neurons of the top layers of V1. Thus, besides the local contrast border information, neurons were found to encode the side to which the border belongs ("border ownership coding"). A majority of these neurons coded border ownership and the local polarity of luminance-chromaticity contrast. The others were insensitive to contrast polarity. Another 20% of the neurons of V2 and V4, and 48% of top layer V1, coded local contrast polarity, but not border ownership. The border ownership-related response differences emerged soon (<25 msec) after the response onset. In V2 and V4, the differences were found to be nearly independent of figure size up to the limit set by the size of our display (21 degrees ). Displays that differed only far outside the conventional receptive field could produce markedly different responses. When tested with more complex displays in which figure-ground cues were varied, some neurons produced invariant border ownership signals, others failed to signal border ownership for some of the displays, but neurons that reversed signals were rare. The influence of visual stimulation far from the receptive field center indicates mechanisms of global context integration. The short latencies and incomplete cue invariance suggest that the border-ownership effect is generated within the visual cortex rather than projected down from higher levels.
Figures






















References
-
- Allman J, Miezin F, McGuinness E. Direction- and velocity-specific responses from beyond the classical receptive field in the middle temporal visual area (MT). Perception. 1985;14:105–126. - PubMed
-
- Anderson BL, Nakayama K. Toward a general theory of stereopsis: binocular matching, occluding contours, and fusion. Psychol Rev. 1994;101:414–445. - PubMed
-
- Baumann R, van der Zwan R, Peterhans E. Figure-ground segregation at contours: a neural mechanism in the visual cortex of the alert monkey. Eur J Neurosci. 1997;9:1290–1303. - PubMed
-
- Bullier J, Nowak LG. Parallel versus serial processing: new vistas on the distributed organization of the visual system. Curr Opin Neurobiol. 1995;5:497–503. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources