

Bacterial SLH domain proteins are non-covalently anchored to the cell surface via a conserved mechanism involving wall polysaccharide pyruvylation
- PMID: 10970841
- PMCID: PMC302060
- DOI: 10.1093/emboj/19.17.4473
Bacterial SLH domain proteins are non-covalently anchored to the cell surface via a conserved mechanism involving wall polysaccharide pyruvylation
Abstract
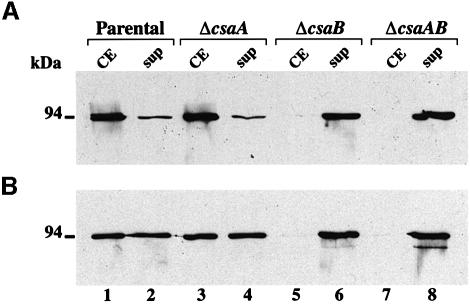
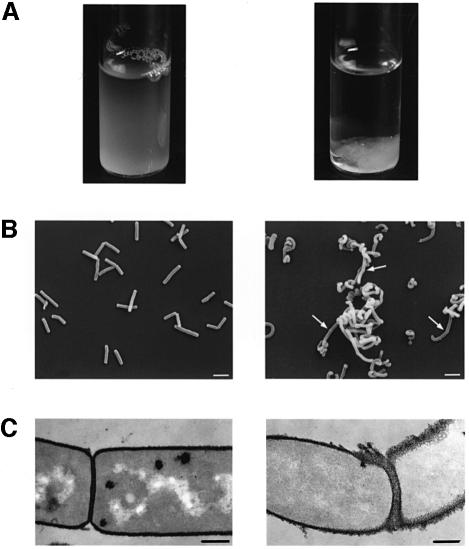
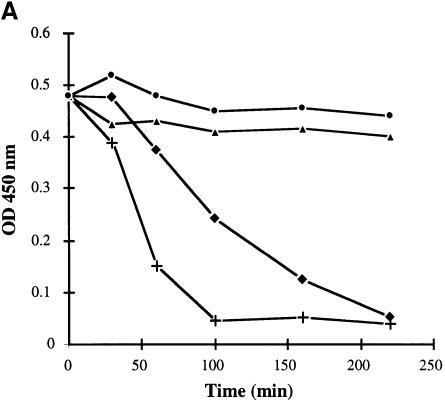
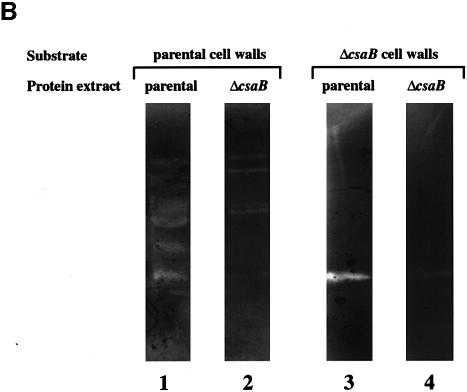
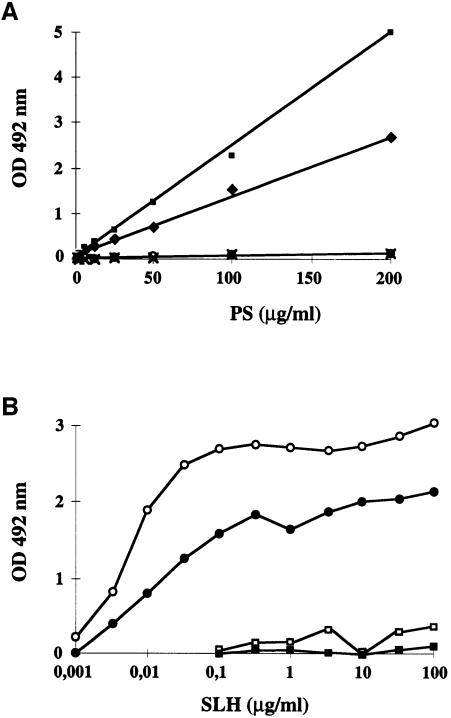
Several bacterial proteins are non-covalently anchored to the cell surface via an S-layer homology (SLH) domain. Previous studies have suggested that this cell surface display mechanism involves a non-covalent interaction between the SLH domain and peptidoglycan-associated polymers. Here we report the characterization of a two-gene operon, csaAB, for cell surface anchoring, in Bacillus anthracis. Its distal open reading frame (csaB) is required for the retention of SLH-containing proteins on the cell wall. Biochemical analysis of cell wall components showed that CsaB was involved in the addition of a pyruvyl group to a peptidoglycan-associated polysaccharide fraction, and that this modification was necessary for binding of the SLH domain. The csaAB operon is present in several bacterial species that synthesize SLH-containing proteins. This observation and the presence of pyruvate in the cell wall of the corresponding bacteria suggest that the mechanism described in this study is widespread among bacteria.
Figures
References
-
- Bray D.F., Bagu,J. and Koegler,P. (1993) Comparison of hexamethyldisilane (HMDS), peldri II, and critical-point drying methods for scanning electron microscopy of biological specimens. Micro. Res. Tech., 26, 489–495. - PubMed
-
- Bugert P. and Geider,K. (1995) Molecular analysis of the ams operon required for expopolysaccharide synthesis of Erwinia amylovora. Mol. Microbiol., 15, 917–933. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
