

Localization of phosphatidylinositol 3-phosphate in yeast and mammalian cells
- PMID: 10970851
- PMCID: PMC302054
- DOI: 10.1093/emboj/19.17.4577
Localization of phosphatidylinositol 3-phosphate in yeast and mammalian cells
Abstract
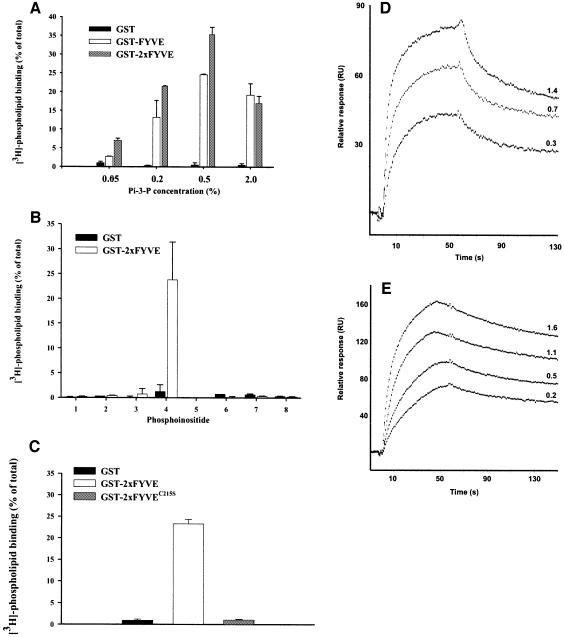
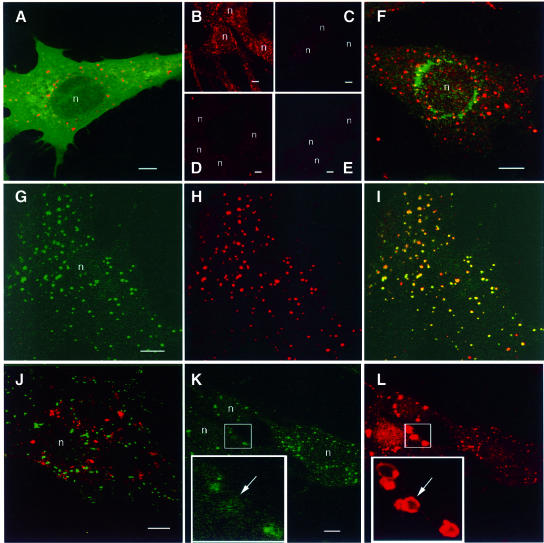
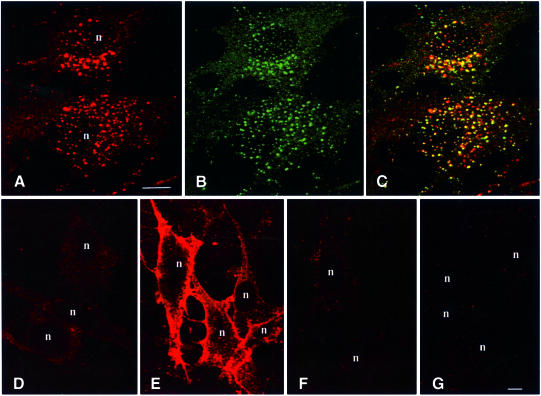
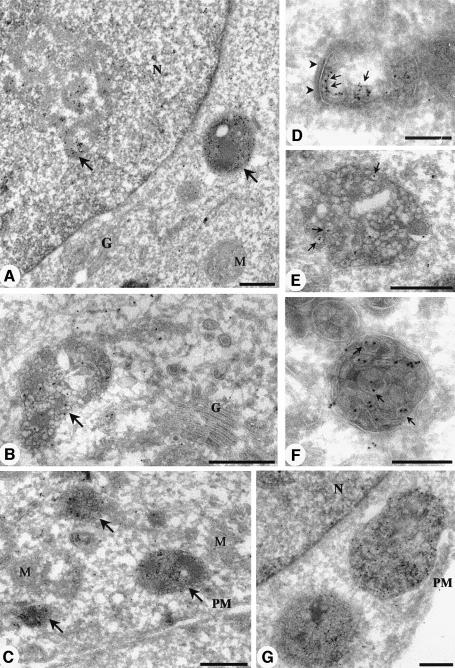
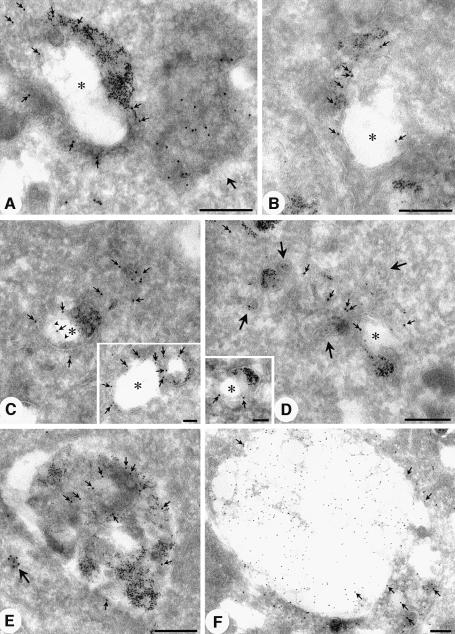
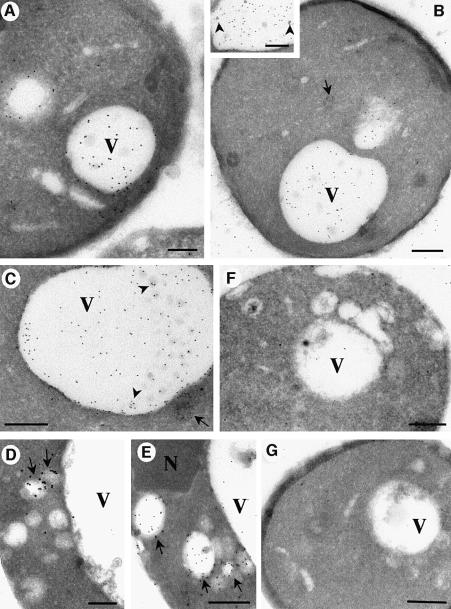
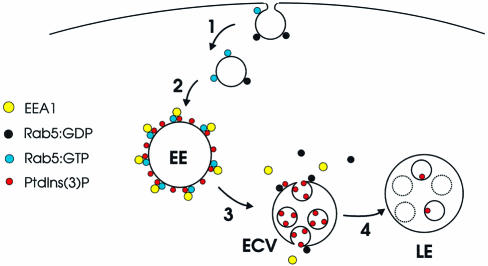
Phosphatidylinositol 3-kinase (PI3K) regulates several vital cellular processes, including signal transduction and membrane trafficking. In order to study the intracellular localization of the PI3K product, phosphatidylinositol 3-phosphate [PI(3)P], we constructed a probe consisting of two PI(3)P-binding FYVE domains. The probe was found to bind specifically, and with high affinity, to PI(3)P both in vitro and in vivo. When expressed in fibroblasts, a tagged probe localized to endosomes, as detected by fluorescence microscopy. Electron microscopy of untransfected fibroblasts showed that PI(3)P is highly enriched on early endosomes and in the internal vesicles of multivesicular endosomes. While yeast cells deficient in PI3K activity (vps15 and vps34 mutants) were not labelled, PI(3)P was found on intralumenal vesicles of endosomes and vacuoles of wild-type yeast. vps27Delta yeast cells, which have impaired endosome to vacuole trafficking, showed a decreased vacuolar labelling and increased endosome labelling. Thus PI(3)P follows a conserved intralumenal degradation pathway, and its generation, accessibility and turnover are likely to play a crucial role in defining the early endosome and the subsequent steps leading to multivesicular endosome formation.
Figures
References
-
- Burd C.G. and Emr,S.D. (1998) Phosphatidylinositol(3)-phosphate signaling mediated by specific binding to RING FYVE domains. Mol. Cell, 2, 157–162. - PubMed
-
- Christoforidis S., Miaczynska,M., Ashman,K., Wilm,M., Zhao,L., Yip,S.-C., Waterfield,M.D., Backer,J.M. and Zerial,M. (1999) Phosphatidylinositol-3-OH kinases are Rab5 effectors. Nature Cell Biol., 1, 249–252. - PubMed
-
- Corvera S., D’Arrigo,A. and Stenmark,H. (1999) Phosphoinositides in membrane traffic. Curr. Opin. Cell Biol., 11, 460–465. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
