

Regulation of origin recognition complex conformation and ATPase activity: differential effects of single-stranded and double-stranded DNA binding
- PMID: 10970868
- PMCID: PMC302069
- DOI: 10.1093/emboj/19.17.4774
Regulation of origin recognition complex conformation and ATPase activity: differential effects of single-stranded and double-stranded DNA binding
Abstract
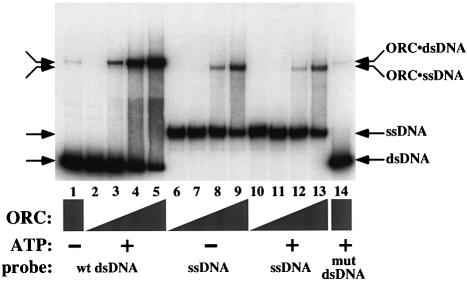
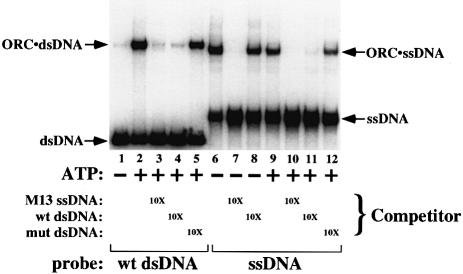
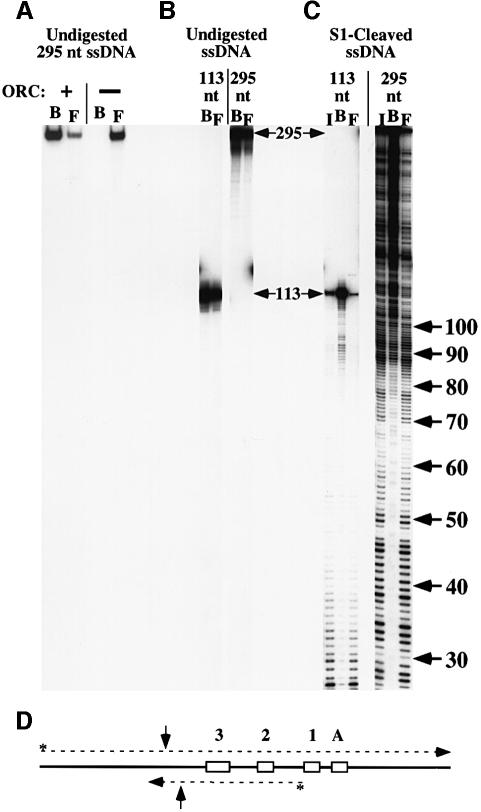
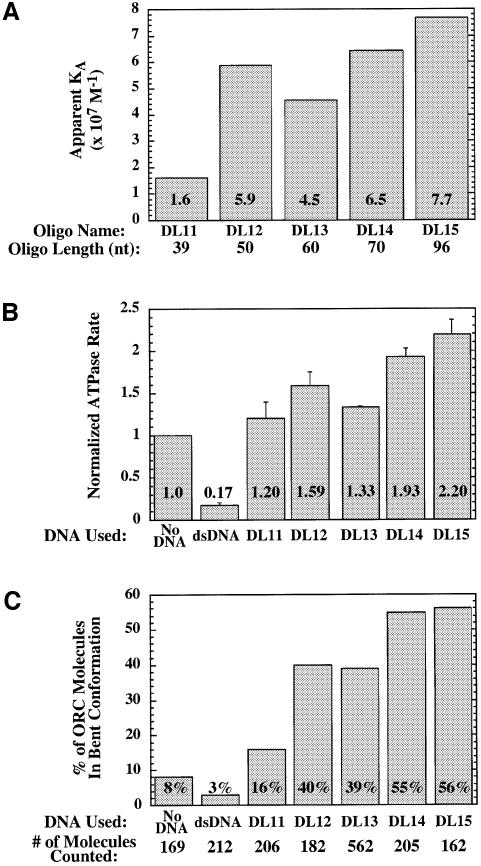
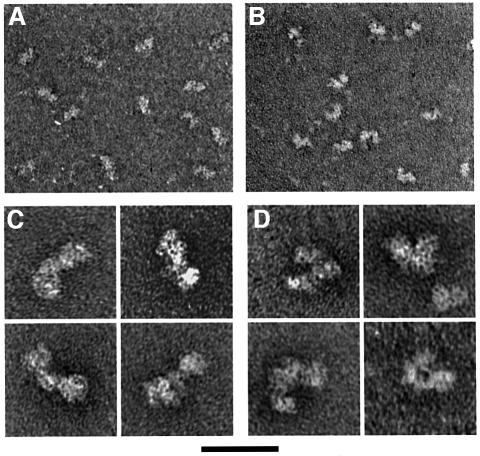
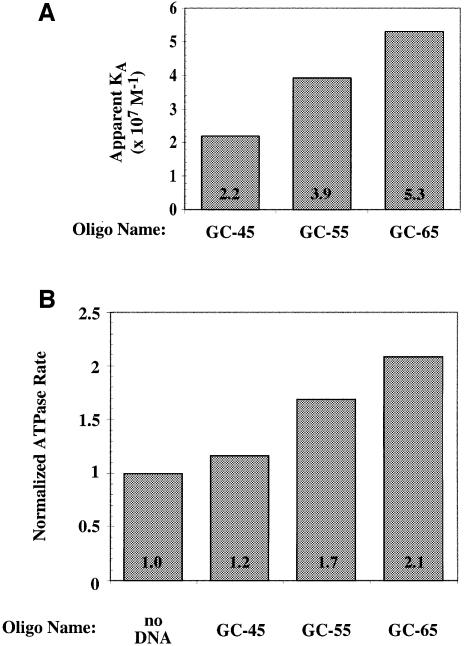
The Saccharomyces cerevisiae origin recognition complex (ORC) is bound to origins of DNA replication throughout the cell cycle and directs the assembly of higher-order protein-DNA complexes during G(1). To examine the fate of ORC when origin DNA is unwound during replication initiation, we determined the effect of single-stranded DNA (ssDNA) on ORC. We show that ORC can bind ssDNA and that ORC bound to ssDNA is distinct from that bound to double-stranded origin DNA. ssDNA stimulated ORC ATPase activity, whereas double-stranded origin DNA inhibited the same activity. Electron microscopy studies revealed two alternative conformations of ORC: an extended conformation stabilized by origin DNA and a bent conformation stabilized by ssDNA. Therefore, ORC appears to exist in two distinct states with respect to its conformation and ATPase activity. Interestingly, the effect of ssDNA on these properties of ORC is correlated with ssDNA length. Since double-stranded origin DNA and ssDNA differentially stabilize these two forms of ORC, we propose that origin unwinding triggers a transition between these alternative states.
Figures
Similar articles
-
Specific basic patch-dependent multimerization of Saccharomyces cerevisiae ORC on single-stranded DNA promotes ATP hydrolysis.Genes Cells. 2019 Sep;24(9):608-618. doi: 10.1111/gtc.12710. Epub 2019 Jul 10. Genes Cells. 2019. PMID: 31233675
-
Coordinate binding of ATP and origin DNA regulates the ATPase activity of the origin recognition complex.Cell. 1997 Feb 21;88(4):493-502. doi: 10.1016/s0092-8674(00)81889-9. Cell. 1997. PMID: 9038340
-
Human origin recognition complex binds preferentially to G-quadruplex-preferable RNA and single-stranded DNA.J Biol Chem. 2013 Oct 18;288(42):30161-30171. doi: 10.1074/jbc.M113.492504. Epub 2013 Sep 3. J Biol Chem. 2013. PMID: 24003239 Free PMC article.
-
Multiple functions of the origin recognition complex.Int Rev Cytol. 2007;256:69-109. doi: 10.1016/S0074-7696(07)56003-1. Int Rev Cytol. 2007. PMID: 17241905 Review.
-
DNA polymerase clamp loaders and DNA recognition.FEBS Lett. 2005 Feb 7;579(4):863-7. doi: 10.1016/j.febslet.2004.11.038. FEBS Lett. 2005. PMID: 15680964 Review.
Cited by
-
Cdc6: a multi-functional molecular switch with critical role in carcinogenesis.Transcription. 2012 May-Jun;3(3):124-9. doi: 10.4161/trns.20301. Transcription. 2012. PMID: 22771947 Free PMC article. Review.
-
Characterization of a novel origin recognition complex-like complex: implications for DNA recognition, cell cycle control, and locus-specific gene amplification.Mol Cell Biol. 2003 Jul;23(14):5005-17. doi: 10.1128/MCB.23.14.5005-5017.2003. Mol Cell Biol. 2003. PMID: 12832485 Free PMC article.
-
Genomewide and biochemical analyses of DNA-binding activity of Cdc6/Orc1 and Mcm proteins in Pyrococcus sp.Nucleic Acids Res. 2007;35(10):3214-22. doi: 10.1093/nar/gkm212. Epub 2007 Apr 22. Nucleic Acids Res. 2007. PMID: 17452353 Free PMC article.
-
ATP bound to the origin recognition complex is important for preRC formation.Proc Natl Acad Sci U S A. 2001 Jul 17;98(15):8361-7. doi: 10.1073/pnas.131006898. Proc Natl Acad Sci U S A. 2001. PMID: 11459976 Free PMC article.
-
RAD51-independent break-induced replication to repair a broken chromosome depends on a distant enhancer site.Genes Dev. 2001 May 1;15(9):1055-60. doi: 10.1101/gad.875901. Genes Dev. 2001. PMID: 11331601 Free PMC article.
References
-
- Aparicio O.M., Weinstein,D.M. and Bell,S.P. (1997) Components and dynamics of DNA replication complexes in S. cerevisiae: redistribution of MCM proteins and Cdc45p during S phase. Cell, 91, 59–69. - PubMed
-
- Ausubel F.M., Brent,R., Kingston,R.E., Moore,D.D., Seidman,J.G., Smith,J.A. and Struhl,K. (1994) Current Protocols in Molecular Biology. John Wiley & Sons, Inc., New York, NY.
-
- Baker T.A. and Bell,S.P. (1998) Polymerases and the replisome: machines within machines. Cell, 92, 295–305. - PubMed
-
- Bell S.P. and Stillman,B. (1992) ATP-dependent recognition of eukaryotic origins of DNA replication by a multiprotein complex. Nature, 357, 128–134. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
