

The Arabidopsis male-sterile mutant, opr3, lacks the 12-oxophytodienoic acid reductase required for jasmonate synthesis
- PMID: 10973494
- PMCID: PMC27075
- DOI: 10.1073/pnas.190264497
The Arabidopsis male-sterile mutant, opr3, lacks the 12-oxophytodienoic acid reductase required for jasmonate synthesis
Abstract
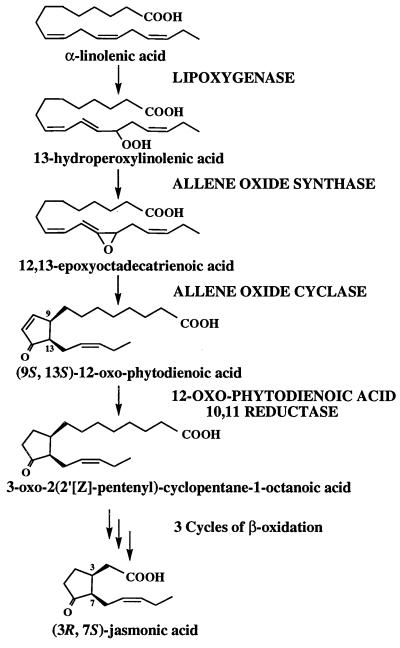
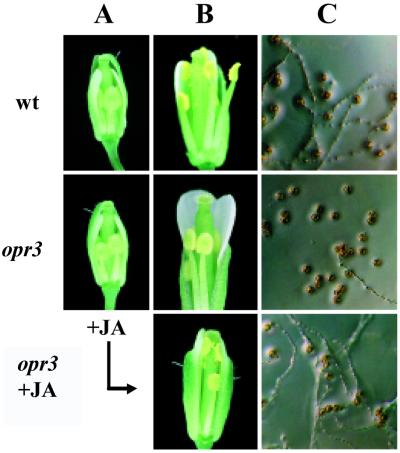
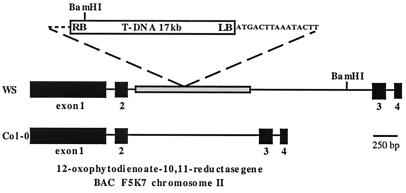
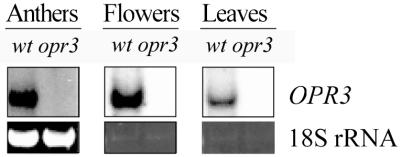
Jasmonic acid (JA) and its precursor 12-oxophytodienoic acid (OPDA) act as plant growth regulators and mediate responses to environmental cues. To investigate the role of these oxylipins in anther and pollen development, we characterized a T-DNA-tagged, male-sterile mutant of Arabidopsis, opr3. The opr3 mutant plants are sterile but can be rendered fertile by exogenous JA but not by OPDA. Cloning of the mutant locus indicates that it encodes an isozyme of 12-oxophytodienoate reductase, designated OPR3. All of the defects in opr3 are alleviated by transformation of the mutant with an OPR3 cDNA. Our results indicate that JA and not OPDA is the signaling molecule that induces and coordinates the elongation of the anther filament, the opening of the stomium at anthesis, and the production of viable pollen. Just as importantly, our data demonstrate that OPR3 is the only isoform of OPR capable of reducing the correct stereoisomer of OPDA to produce JA required for male gametophyte development.
Figures

References
-
- Ryan C A. Annu Rev Plant Physiol Plant Mol Biol. 1991;42:651–674.
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
