

A myosin I is involved in membrane recycling from early endosomes
- PMID: 10973992
- PMCID: PMC2175260
- DOI: 10.1083/jcb.150.5.1013
A myosin I is involved in membrane recycling from early endosomes
Abstract
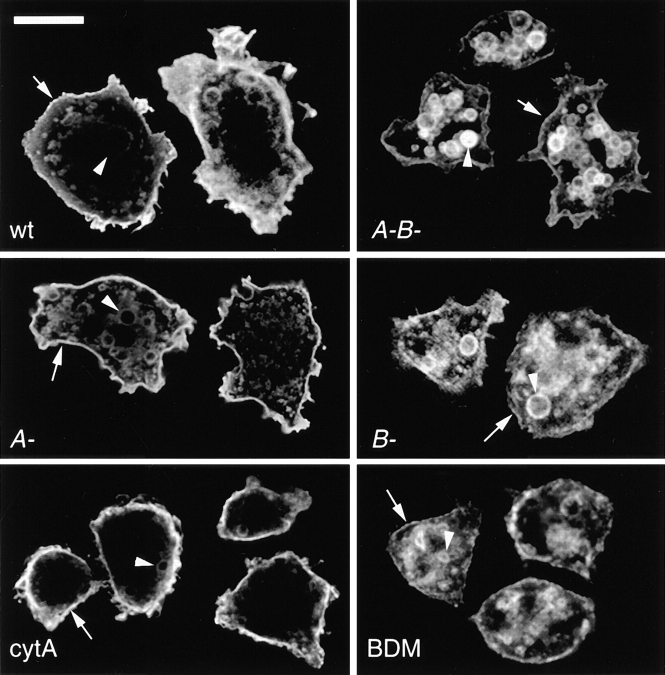
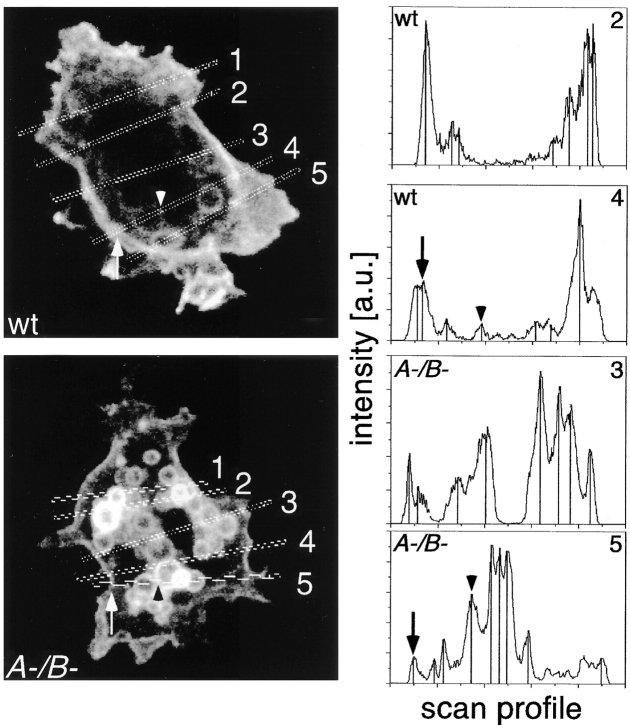
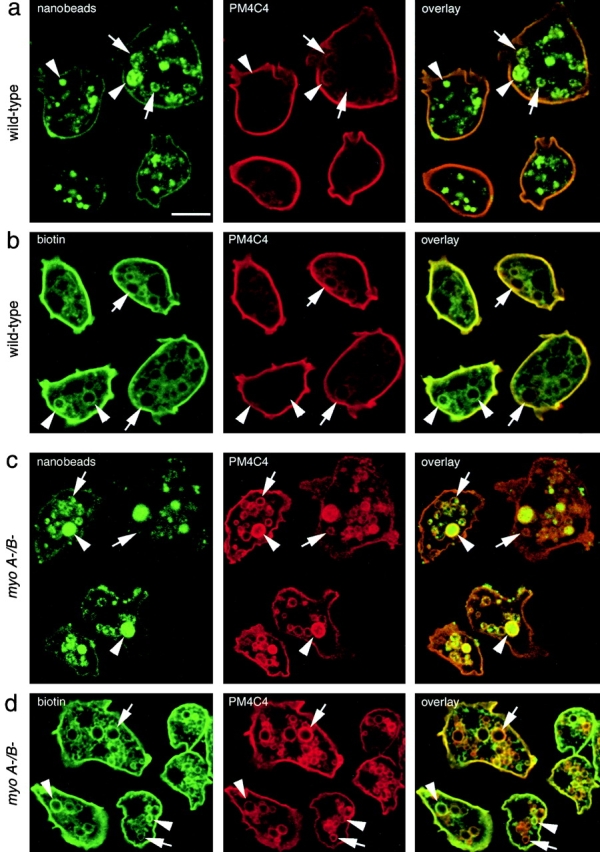
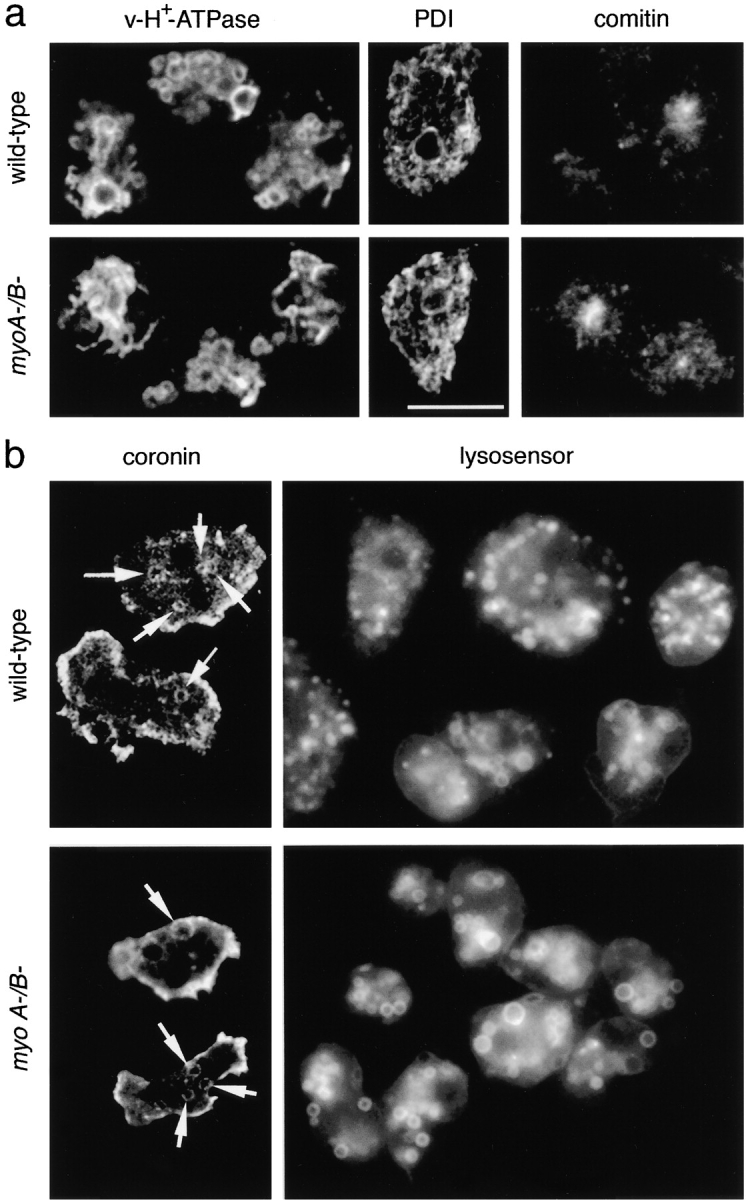
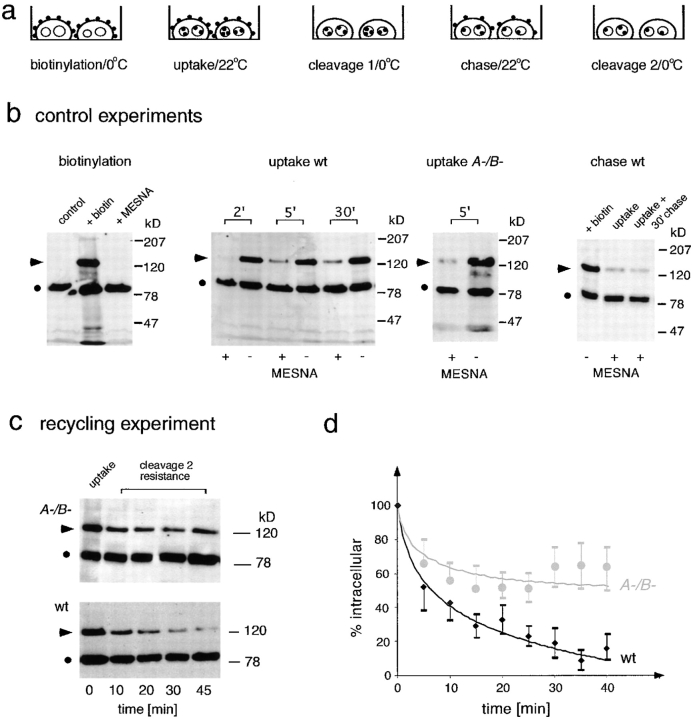
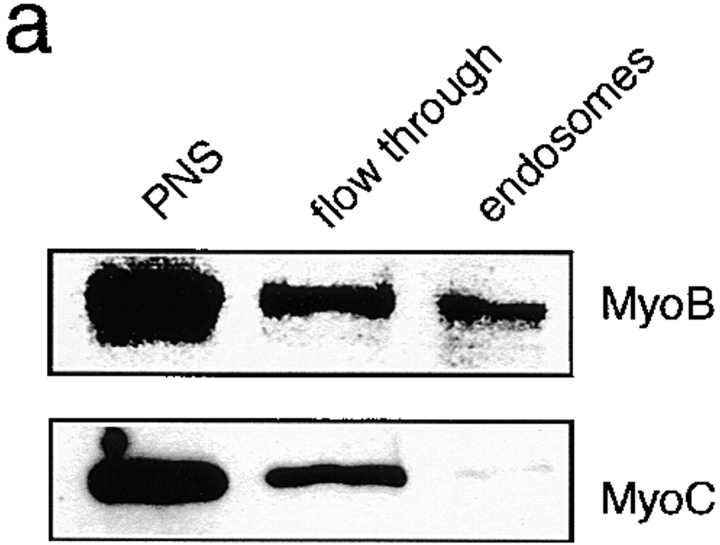
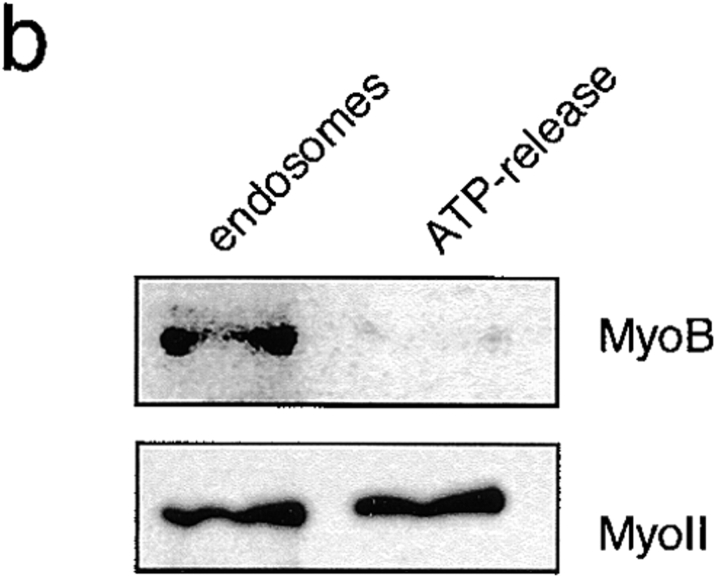
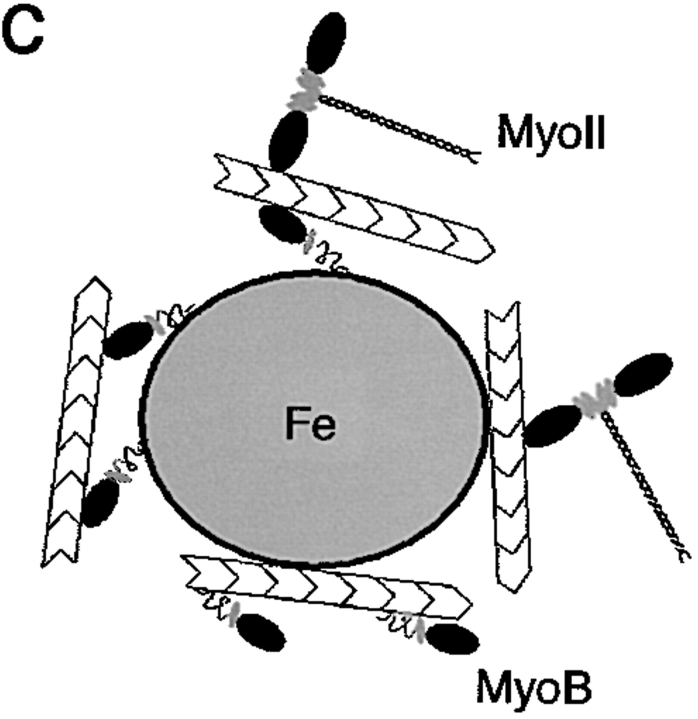
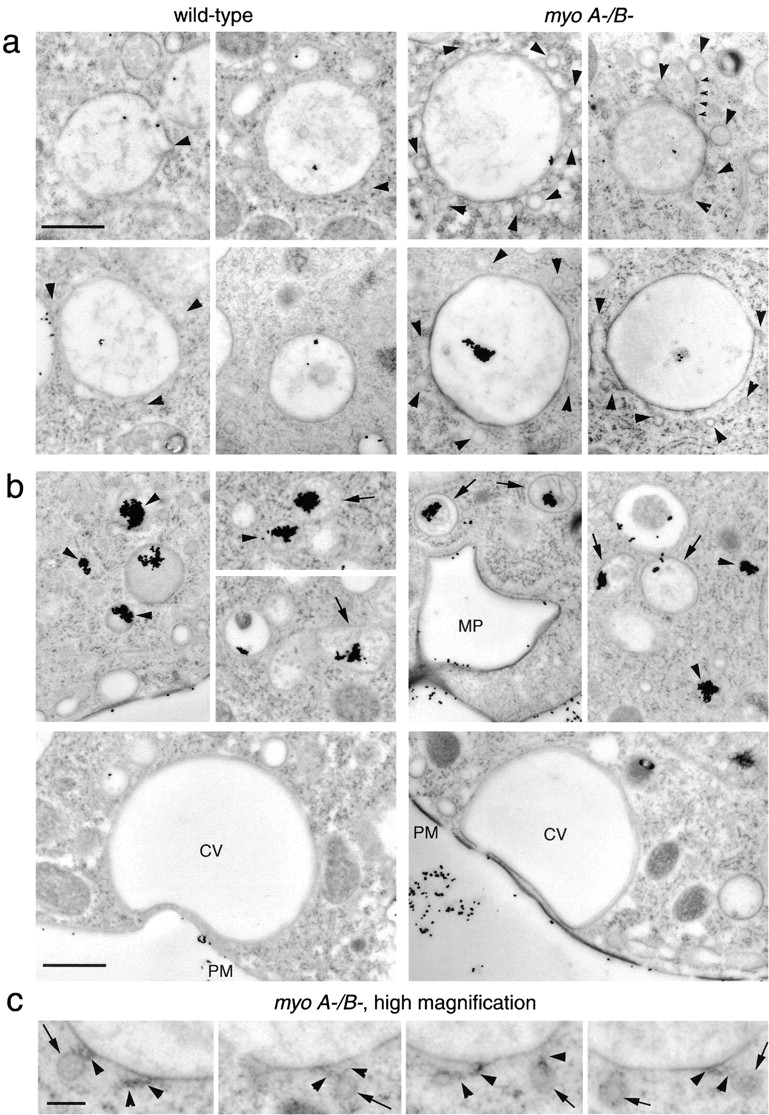
Geometry-based mechanisms have been proposed to account for the sorting of membranes and fluid phase in the endocytic pathway, yet little is known about the involvement of the actin-myosin cytoskeleton. Here, we demonstrate that Dictyostelium discoideum myosin IB functions in the recycling of plasma membrane components from endosomes back to the cell surface. Cells lacking MyoB (myoA(-)/B(-), and myoB(-) cells) and wild-type cells treated with the myosin inhibitor butanedione monoxime accumulated a plasma membrane marker and biotinylated surface proteins on intracellular endocytic vacuoles. An assay based on reversible biotinylation of plasma membrane proteins demonstrated that recycling of membrane components is severely impaired in myoA/B null cells. In addition, MyoB was specifically found on magnetically purified early pinosomes. Using a rapid-freezing cryoelectron microscopy method, we observed an increased number of small vesicles tethered to relatively early endocytic vacuoles in myoA(-)/B(-) cells, but not to later endosomes and lysosomes. This accumulation of vesicles suggests that the defects in membrane recycling result from a disordered morphology of the sorting compartment.
Figures
Similar articles
-
Examination of the endosomal and lysosomal pathways in Dictyostelium discoideum myosin I mutants.J Cell Sci. 1996 Mar;109 ( Pt 3):663-73. doi: 10.1242/jcs.109.3.663. J Cell Sci. 1996. PMID: 8907711
-
Transport from late endosomes to lysosomes, but not sorting of integral membrane proteins in endosomes, depends on the vacuolar proton pump.J Cell Biol. 1995 Aug;130(4):821-34. doi: 10.1083/jcb.130.4.821. J Cell Biol. 1995. PMID: 7642700 Free PMC article.
-
Maturation of early endosomes and vesicular traffic to lysosomes in relation to membrane recycling.J Cell Sci. 1995 Apr;108 ( Pt 4):1791-803. doi: 10.1242/jcs.108.4.1791. J Cell Sci. 1995. PMID: 7542261
-
Endocytic traffic in polarized epithelial cells: role of the actin and microtubule cytoskeleton.Traffic. 2001 Mar;2(3):149-59. doi: 10.1034/j.1600-0854.2001.020301.x. Traffic. 2001. PMID: 11260520 Review.
-
Endocytosis and the actin cytoskeleton in Dictyostelium discoideum.Int Rev Cell Mol Biol. 2008;267:343-97. doi: 10.1016/S1937-6448(08)00633-3. Int Rev Cell Mol Biol. 2008. PMID: 18544503 Review.
Cited by
-
High-resolution dissection of phagosome maturation reveals distinct membrane trafficking phases.Mol Biol Cell. 2002 Oct;13(10):3508-20. doi: 10.1091/mbc.e02-04-0206. Mol Biol Cell. 2002. PMID: 12388753 Free PMC article.
-
Dictyostelium LvsB mutants model the lysosomal defects associated with Chediak-Higashi syndrome.Mol Biol Cell. 2002 Feb;13(2):656-69. doi: 10.1091/mbc.01-09-0454. Mol Biol Cell. 2002. PMID: 11854420 Free PMC article.
-
Molecular basis of dynamic relocalization of Dictyostelium myosin IB.J Biol Chem. 2012 Apr 27;287(18):14923-36. doi: 10.1074/jbc.M111.318667. Epub 2012 Feb 24. J Biol Chem. 2012. PMID: 22367211 Free PMC article.
-
Actin filaments and myosin I alpha cooperate with microtubules for the movement of lysosomes.Mol Biol Cell. 2001 Dec;12(12):4013-29. doi: 10.1091/mbc.12.12.4013. Mol Biol Cell. 2001. PMID: 11739797 Free PMC article.
-
Calmodulin dissociation regulates Myo5 recruitment and function at endocytic sites.EMBO J. 2010 Sep 1;29(17):2899-914. doi: 10.1038/emboj.2010.159. Epub 2010 Jul 20. EMBO J. 2010. PMID: 20647997 Free PMC article.
References
-
- Abercrombie M., Heaysman J.E.M., Pegrum S.M. The locomotion of fibroblasts in culture. 3. Movement of particles on the dorsal surface of the leading lamella. Exp. Cell Res. 1970;62:389–398. - PubMed
-
- Adessi C., Chapel A., Vincon M., Rabilloud T., Klein G., Satre M., Garin J. Identification of major proteins associated with Dictyostelium discoideum endocytic vesicles. J. Cell Sci. 1995;108:3331–3337. - PubMed
-
- Aubry L., Klein G., Martiel J.L., Satre M. Kinetics of endosomal pH evolution in Dictyostelium discoideum amoebae. Study by fluorescence spectroscopy. J. Cell Sci. 1993;105:861–866. - PubMed
-
- Aubry L., Klein G., Martiel J.L., Satre M. Fluid-phase endocytosis in the amoebae of the cellular slime mould Dictyostelium discoideummathematical modelling of kinetics and pH evolution. J. Theor. Biol. 1997;184:89–98.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
