

doi: 10.1084/jem.192.5.659.
Cloning of the murine thymic stromal lymphopoietin (TSLP) receptor: Formation of a functional heteromeric complex requires interleukin 7 receptor
Affiliations
- PMID: 10974032
- PMCID: PMC2193276
- DOI: 10.1084/jem.192.5.659
Item in Clipboard
Cloning of the murine thymic stromal lymphopoietin (TSLP) receptor: Formation of a functional heteromeric complex requires interleukin 7 receptor
J Exp Med.
.
Abstract
The cellular receptor for murine thymic stromal lymphopoietin (TSLP) was detected in a variety of murine, but not human myelomonocytic cell lines by radioligand binding. cDNA clones encoding the receptor were isolated from a murine T helper cell cDNA library. TSLP receptor (TSLPR) is a member of the hematopoietin receptor family. Transfection of TSLPR cDNA resulted in only low affinity binding. Cotransfection of the interleukin 7 (IL-7)Ralpha chain cDNA resulted in conversion to high affinity binding. TSLP did not activate cells from IL-7Ralpha(-/)- mice, but did activate cells from gammac(-/)- mice. Thus, the functional TSLPR requires the IL-7Ralpha chain, but not the gammac chain for signaling.
Figures
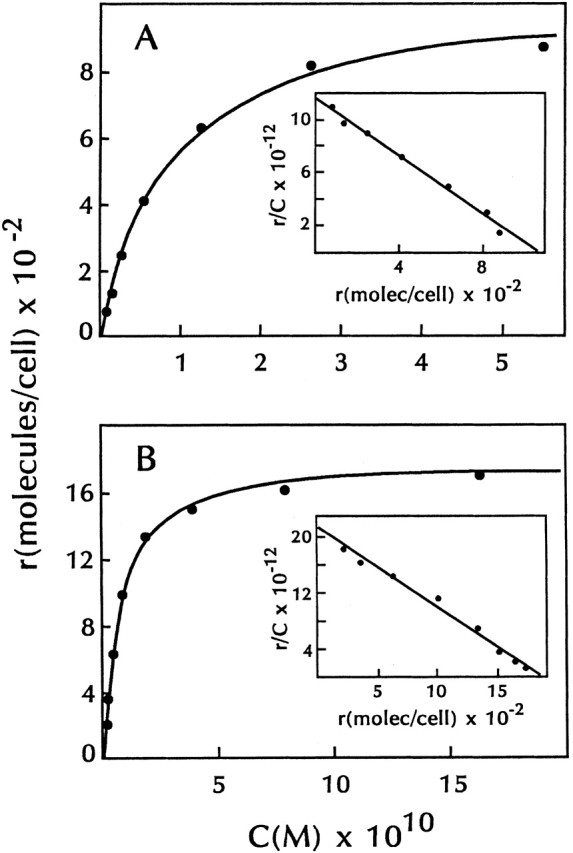
Equilibrium binding of 125I-TSLP to lymphoid cell lines. 70Z/3 cells (A) and anti-CD3–stimulated 7B9 cells (B) were incubated with various concentrations of 125I-TSLP for 30 min at 37°C. Binding was assayed as described in Materials and Methods. All data were corrected for nonspecific binding. Insets show Scatchard representations of specific binding from each panel.
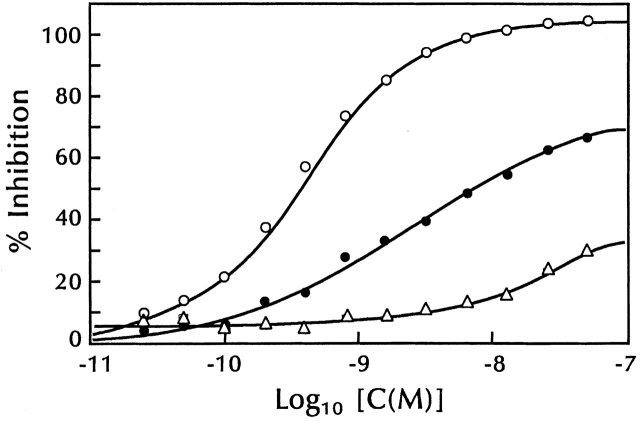
Inhibition of 125I-TSLP binding by unlabeled TSLP and IL-7. 70Z/3 cells (5 × 107 cells) were incubated with 125I-TSLP (2.6 × 10−10 M) and varying concentrations of unlabeled TSLP (○), murine IL-7 (•), and human IL-7 (▵) for 30 min at 37°C. Binding was assayed as described in Materials and Methods. The continuous curves were calculated from either a one-site (TSLP) or two-site (murine and human IL-7) competitive inhibition equation using a K
a value for 125I-TSLP of 8.8 × 109 M−1. All data were corrected for nonspecific binding.

Murine TSLPR cDNA. (A) The cDNA and predicted amino acid sequence of the murine TSLPR clone 7a. Signal peptide cleavage is shown by the arrow as occurring after Ala 21, although cleavage after Ala 15 would be nearly as preferred (reference 37). The transmembrane region is underlined; asterisks mark the cysteines conserved in hematopoietin family members; a double underline indicates the sequence WTAVT, present here in place of the more typical WSXWS; the box 1 sequence of Murakami et al. is boxed (reference 28). A second functional cDNA clone isolated in the expression screen, clone 19, extends from nucleotide 80 through the polyA stretch at the 3′ end. The initiating Met-Ala-Trp sequence at the NH2 terminus of clone 7a is replaced in clone 19 by Met-Gly, encoded in the adapter used to insert the cDNA into the vector. (B) Alignment of the membrane-proximal segment of the murine TSLPR with membrane-proximal segments of the murine erythropoietin receptor (EpoR), the murine IL-2 receptor β chain, and the murine IL-9 receptor, with regions of amino acid conservation highlighted.

Murine TSLPR cDNA. (A) The cDNA and predicted amino acid sequence of the murine TSLPR clone 7a. Signal peptide cleavage is shown by the arrow as occurring after Ala 21, although cleavage after Ala 15 would be nearly as preferred (reference 37). The transmembrane region is underlined; asterisks mark the cysteines conserved in hematopoietin family members; a double underline indicates the sequence WTAVT, present here in place of the more typical WSXWS; the box 1 sequence of Murakami et al. is boxed (reference 28). A second functional cDNA clone isolated in the expression screen, clone 19, extends from nucleotide 80 through the polyA stretch at the 3′ end. The initiating Met-Ala-Trp sequence at the NH2 terminus of clone 7a is replaced in clone 19 by Met-Gly, encoded in the adapter used to insert the cDNA into the vector. (B) Alignment of the membrane-proximal segment of the murine TSLPR with membrane-proximal segments of the murine erythropoietin receptor (EpoR), the murine IL-2 receptor β chain, and the murine IL-9 receptor, with regions of amino acid conservation highlighted.
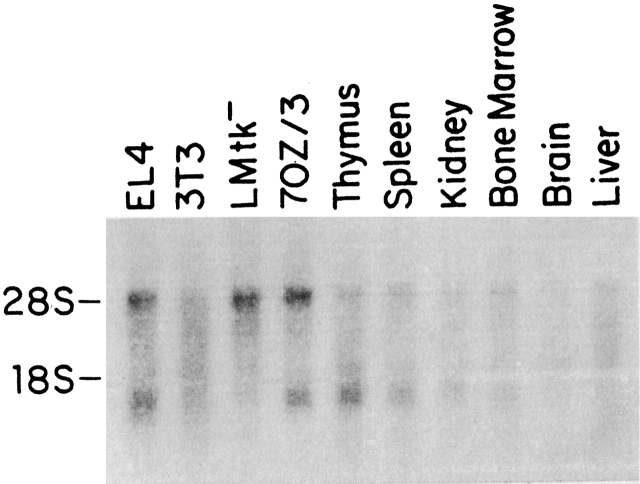
Northern blot analysis of TSLPR mRNA. PolyA+ RNA from various cell lines and tissues was hybridized to an antisense TSLPR riboprobe as described in Materials and Methods. The band at 28S reflects the cross-hybridization of the TSLPR probe to residual rRNA present in the samples.
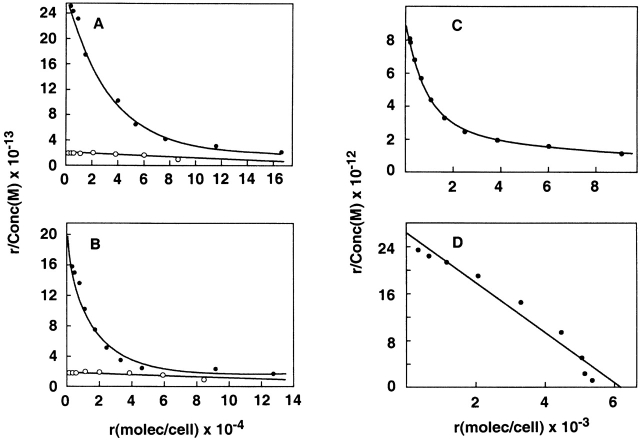
Binding characteristics of TSLPR with the IL-7Rα chain. (A and B) Binding of 125I-muTSLP to CV-1/EBNA cells transfected with murine TSLPR with (•) or without (○) murine IL-7Rα (A) or human IL-7Rα (B). (C and D) Binding of 125I–muIL-7 (C) or 125I-muTSLP (D) to BAF/BO3 cells transfected with the murine IL-7Rα. In all cases binding was done for 30 min at 37°C and assayed as described in Materials and Methods.
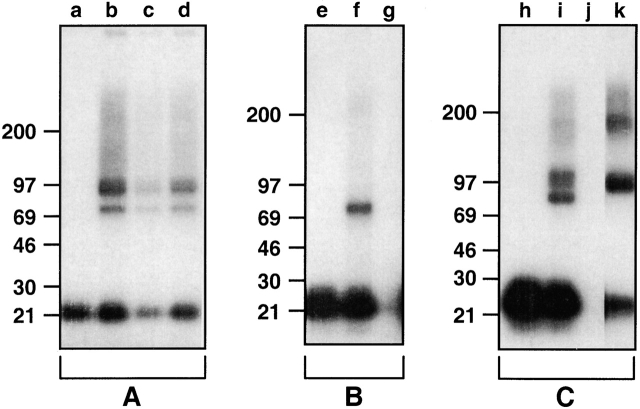
Characterization of the TSLPR by affinity cross-linking. (A) 70Z/3 cells were incubated with 125I-TSLP in the absence of unlabeled competitor (a and b), or in the presence of unlabeled murine IL-7 (c) or human IL-7 (d) and then treated with (b–d) or without (a) cross-linker. (B) CV-1 cells transfected with TSLPR clone 7a were incubated with 125I-TSLP in the absence of unlabeled competitor (e and f) or in the presence of unlabeled TSLP (g) and then treated with (f and g) or without (e) cross-linker. (C) BAF/BO3 cells transfected with muIL-7Rα were incubated with 125I-TSLP in the absence of unlabeled competitor (h and i) or in the presence of unlabeled TSLP (j), or with 125I–IL-7 (k) and then treated with (i–k) or without (h) cross-linker. Cross-linking was performed as described in Materials and Methods and samples were run on SDS-PAGE under reducing conditions using 8–16% gradient gels and analyzed by autoradiography.
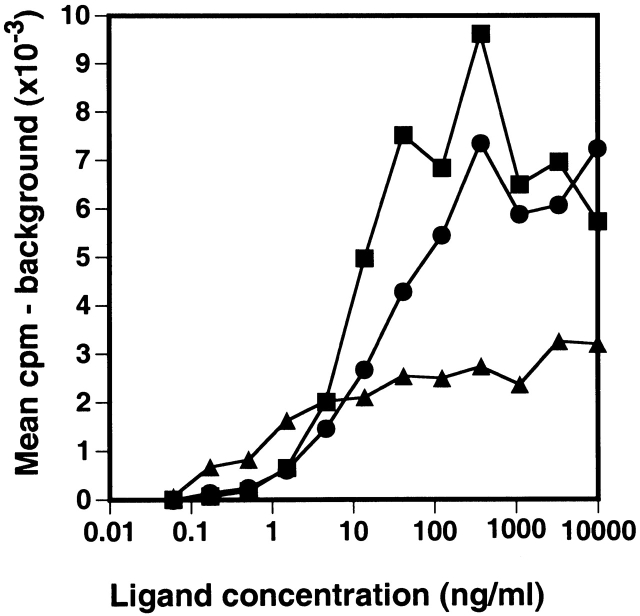
Proliferative response of BAF/BO3 cells transfected with the muIL7Rα chain. BAF/BO3 cells expressing murine IL-7Rα chain (2 × 105 cells/ml) were cultured in medium alone, murine IL-3 (▪), human IL-7(•), or murine TSLP (▴) for 2 d. Cells were then pulsed for 5 h with 3H-TdR and harvested. The background cpm (cells in medium alone) averaged 184 cpm.
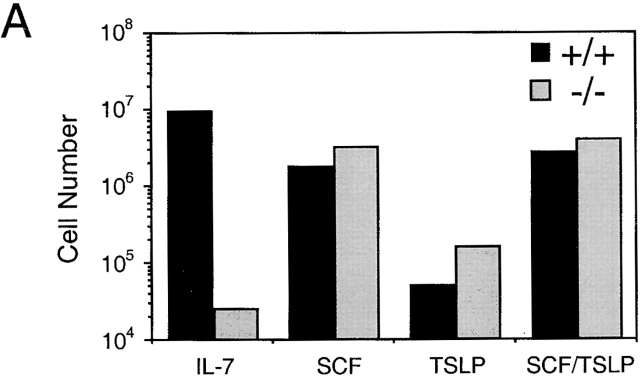
Analysis of TSLP activity in Whitlock-Witte cultures derived from wild-type or IL-7Rα−/− mice. (A) Livers from IL-7Rα−/− or wild-type newborn mice were grown under Whitlock-Witte culture conditions for 9 d in the presence of either IL-7 (10 ng/ml), SCF (1 μg/ml), TSLP (100 ng/ml), or SCF plus TSLP. Enumeration of nonadherent cells was determined by trypan blue exclusion. (B) Immunofluorescent profiles of nonadherent cells isolated from IL-7Rα−/− and wild-type mice. Lymphoid and myeloid gates were established by the forward versus side light scatter profiles of cells grown in IL-7 alone or SCF alone, respectively. Myeloid gated cells were analyzed for Mac-1 and Gr-1 expression after growth in SCF (gray histogram) or SCF plus TSLP (white histogram). Lymphoid gated cells were analyzed for B220 expression after growth in IL-7 (hatched histogram) or SCF plus TSLP (white histogram). Black histogram represents IgG1 isotype control staining of cells grown in IL-7.
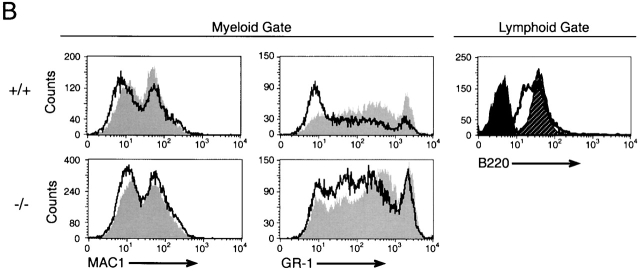
Analysis of TSLP activity in Whitlock-Witte cultures derived from wild-type or IL-7Rα−/− mice. (A) Livers from IL-7Rα−/− or wild-type newborn mice were grown under Whitlock-Witte culture conditions for 9 d in the presence of either IL-7 (10 ng/ml), SCF (1 μg/ml), TSLP (100 ng/ml), or SCF plus TSLP. Enumeration of nonadherent cells was determined by trypan blue exclusion. (B) Immunofluorescent profiles of nonadherent cells isolated from IL-7Rα−/− and wild-type mice. Lymphoid and myeloid gates were established by the forward versus side light scatter profiles of cells grown in IL-7 alone or SCF alone, respectively. Myeloid gated cells were analyzed for Mac-1 and Gr-1 expression after growth in SCF (gray histogram) or SCF plus TSLP (white histogram). Lymphoid gated cells were analyzed for B220 expression after growth in IL-7 (hatched histogram) or SCF plus TSLP (white histogram). Black histogram represents IgG1 isotype control staining of cells grown in IL-7.
References
-
- Komschlies K.L., Grzegorzewski K.J., Wiltrout R.H. Diverse immunological and hematological effects of interleukin-7implications for clinical application. J. Leukoc. Biol. 1995;58:623–633. - PubMed
-
- Di Santo J.P., Kühn R., Müller W. Common cytokine receptor γ chain (γc)-dependent cytokinesunderstanding in vivo functions by gene targeting. Immunol. Rev. 1995;148:19–34. - PubMed
-
- Ihle J.N., Witthuhn B.A., Quelle F.W., Yamamoto K., Silvennoinen O. Signaling through the hematopoietic cytokine receptors. Annu. Rev. Immunol. 1995;13:369–398. - PubMed
-
- Chomarat P., Banchereau J. An update on Interleukin-4 and its receptor. Eur. Cytokine Netw. 1997;8:333–344. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
