

Programmed death in bacteria
- PMID: 10974124
- PMCID: PMC99002
- DOI: 10.1128/MMBR.64.3.503-514.2000
Programmed death in bacteria
Abstract
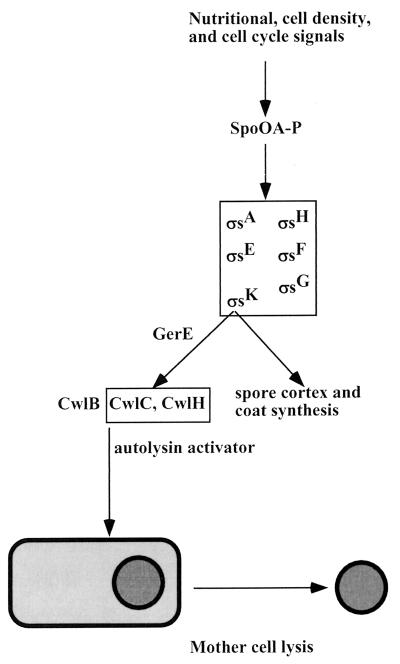
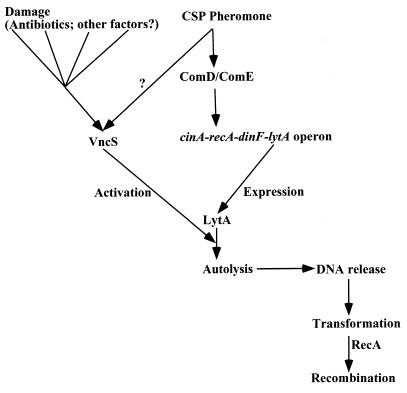
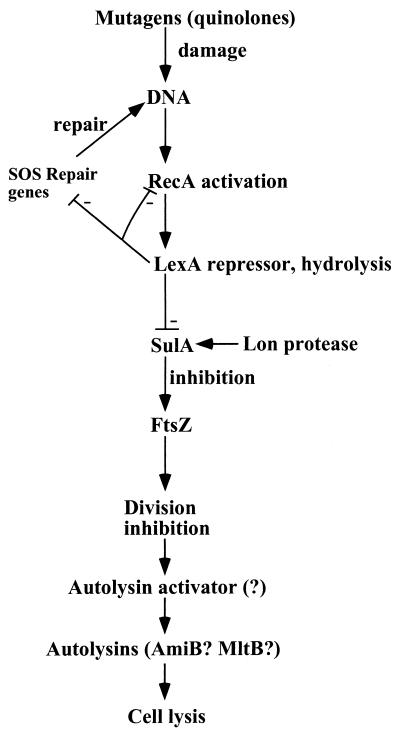
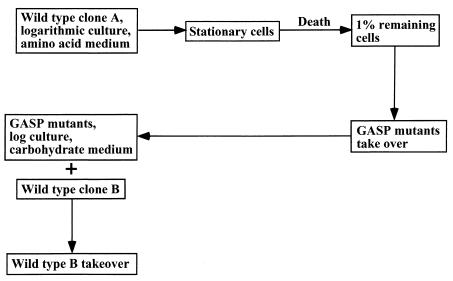
Programmed cell death (PCD) in bacteria plays an important role in developmental processes, such as lysis of the mother cell during sporulation of Bacillus subtilis and lysis of vegetative cells in fruiting body formation of Myxococcus xanthus. The signal transduction pathway leading to autolysis of the mother cell includes the terminal sporulation sigma factor Esigma(K), which induces the synthesis of autolysins CwlC and CwlH. An activator of autolysin in this and other PCD processes is yet to be identified. Autolysis plays a role in genetic exchange in Streptococcus pneumoniae, and the gene for the major autolysin, lytA, is located in the same operon with recA. DNA from lysed cells is picked up by their neighbors and recombined into the chromosome by RecA. LytA requires an unknown activator controlled by a sensory kinase, VncS. Deletion of vncS inhibits autolysis and also decreases killing by unrelated antibiotics. This observation suggests that PCD in bacteria serves to eliminate damaged cells, similar to apoptosis of defective cells in metazoa. The presence of genes affecting survival without changing growth sensitivity to antibiotics (vncS, lytA, hipAB, sulA, and mar) indicates that bacteria are able to control their fate. Elimination of defective cells could limit the spread of a viral infection and donate nutrients to healthy kin cells. An altruistic suicide would be challenged by the appearance of asocial mutants without PCD and by the possibility of maladaptive total suicide in response to a uniformly present lethal factor or nutrient depletion. It is proposed that a low rate of mutation serves to decrease the probability that asocial mutants without PCD will take over the population. It is suggested that PCD is disabled in persistors, rare cells that are resistant to killing, to ensure population survival. It is suggested that lack of nutrients leads to the stringent response that suppresses PCD, producing a state of tolerance to antibiotics, allowing cells to discriminate between nutrient deprivation and unrepairable damage. High levels of persistors are apparently responsible for the extraordinary survival properties of bacterial biofilms, and genes affecting persistence appear to be promising targets for development of drugs aimed at eradicating recalcitrant infections. PCD in unicellular eukaryotes is also considered, including aging in Saccharomyces cerevisiae. Apoptosis-like elimination of defective cells in S. cerevisiae and protozoa suggests that all unicellular life forms evolved altruistic programmed death that serves a variety of useful functions.
Figures
References
-
- Alloing G, Martin B, Granadel C, Claverys J P. Development of competence in Streptococcus pneumoniae pheromone autoinduction and control of quorum sensing by the oligopeptide permease. Mol Microbiol. 1998;29:75–83. - PubMed
-
- Aravind L, Dixit V M, Koonin E V. The domains of death: evolution of the apoptosis machinery. Trends Biochem Sci. 1999;24:47–53. - PubMed
-
- Ashby M J, Neale J E, Knott S J, Critchley I A. Effect of antibiotics on non-growing planktonic cells and biofilms of Escherichia coli. J Antimicrob Chemother. 1994;33:443–452. - PubMed
-
- Beckman K B, Ames B N. The free radical theory of aging matures. Physiol Rev. 1998;78:547–581. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
