

Functional loss of ABCA1 in mice causes severe placental malformation, aberrant lipid distribution, and kidney glomerulonephritis as well as high-density lipoprotein cholesterol deficiency
- PMID: 10980140
- PMCID: PMC1885686
- DOI: 10.1016/S0002-9440(10)64614-7
Functional loss of ABCA1 in mice causes severe placental malformation, aberrant lipid distribution, and kidney glomerulonephritis as well as high-density lipoprotein cholesterol deficiency
Abstract
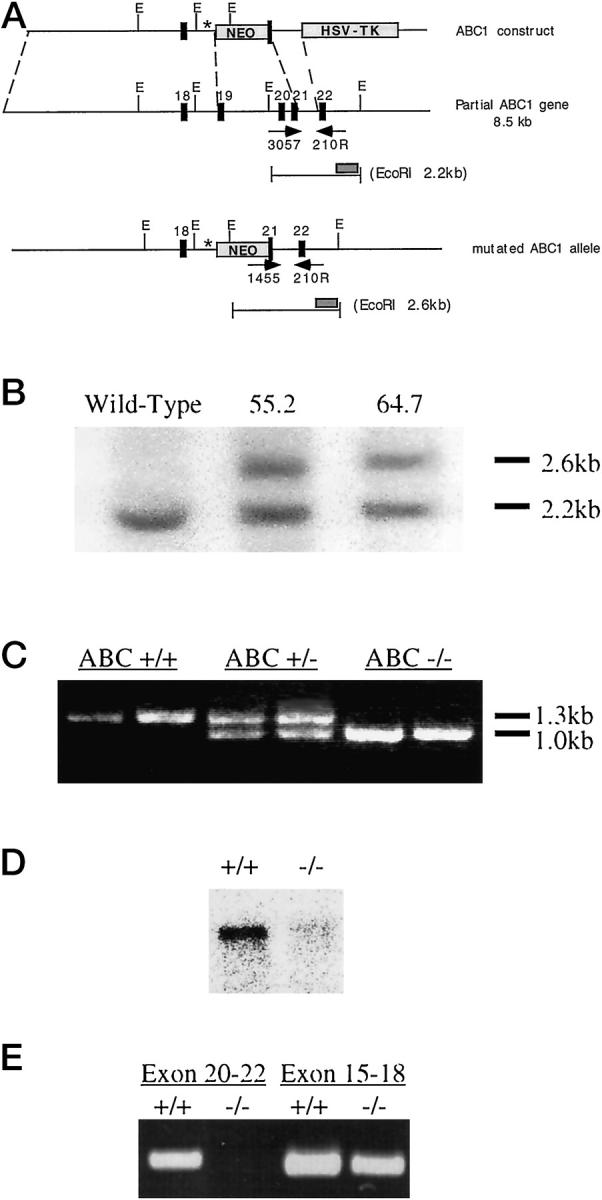
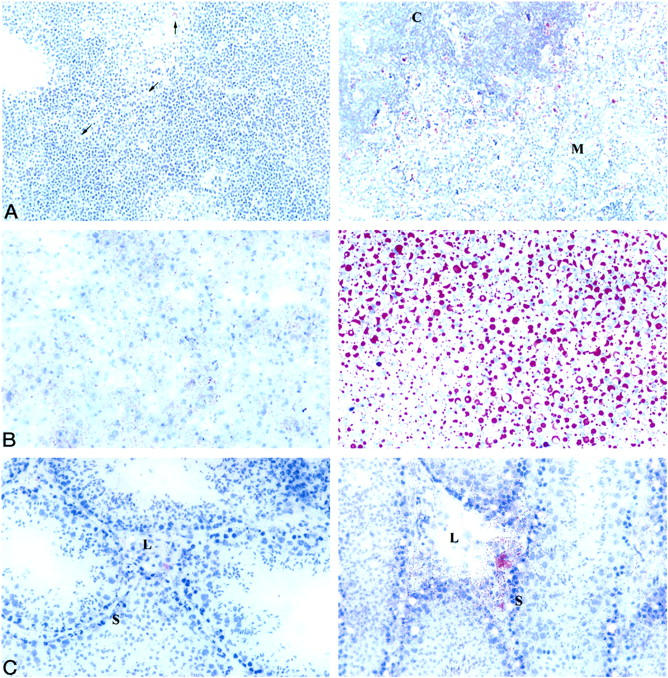
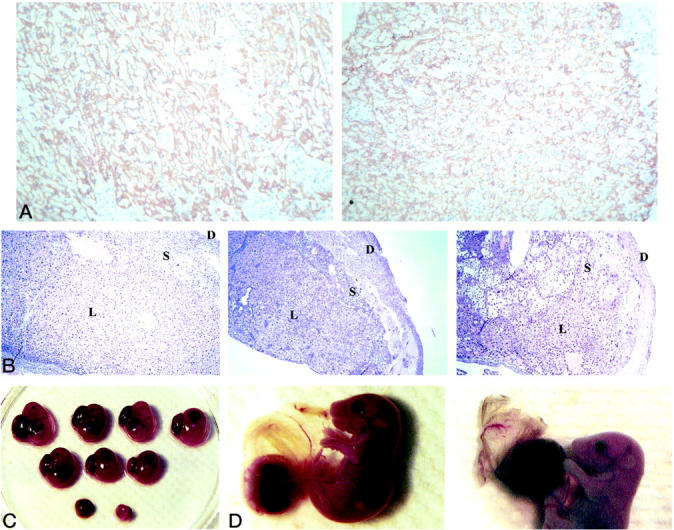
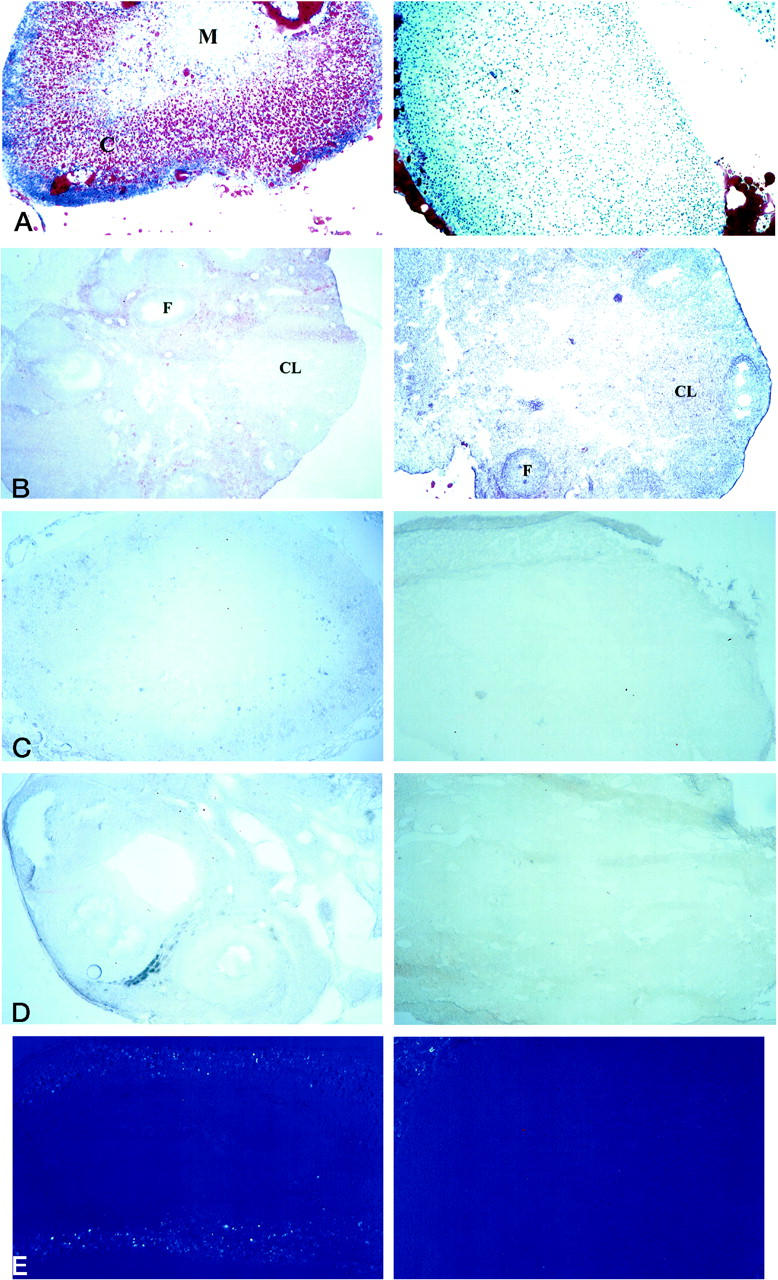
Tangier disease (TD) and familial HDL deficiency (FHA) have recently been linked to mutations in the human ATP-binding cassette transporter 1 (hABCA1), a member of the ABC superfamily. Both diseases are characterized by the lowering or lack of high-density lipoprotein cholesterol (HDL-C) and low serum cholesterol. The murine ABCA1-/- phenotype corroborates the human TD linkage to ABCA1. Similar to TD in humans, HDL-C is virtually absent in ABCA1-/- mice accompanied by a reduction in serum cholesterol and lipid deposition in various tissues. In addition, the placenta of ABCA1-/- mice is malformed, resulting in severe embryo growth retardation, fetal loss, and neonatal death. The basis for these defects appears to be altered steroidogenesis, a direct result of the lack of HDL-C. By 6 months of age, ABCA1-/- animals develop membranoproliferative glomerulonephritis due to deposition of immunocomplexes followed by cardiomegaly with ventricular dilation and hypertrophy, ultimately succumbing to congestive heart failure. This murine model of TD will be very useful in the study of lipid metabolism, renal inflammation, and cardiovascular disease and may reveal previously unsuspected relationships between them.
Figures




Similar articles
-
Decreased cellular cholesterol efflux is a common cause of familial hypoalphalipoproteinemia: role of the ABCA1 gene mutations.Atherosclerosis. 2000 Oct;152(2):457-68. doi: 10.1016/s0021-9150(99)00498-0. Atherosclerosis. 2000. PMID: 10998475
-
The ATP binding cassette transporter A1 (ABCA1) modulates the development of aortic atherosclerosis in C57BL/6 and apoE-knockout mice.Proc Natl Acad Sci U S A. 2002 Jan 8;99(1):407-12. doi: 10.1073/pnas.012587699. Epub 2001 Dec 18. Proc Natl Acad Sci U S A. 2002. PMID: 11752403 Free PMC article.
-
Novel mutations of ABCA1 transporter in patients with Tangier disease and familial HDL deficiency.Mol Genet Metab. 2012 Nov;107(3):534-41. doi: 10.1016/j.ymgme.2012.08.005. Epub 2012 Aug 18. Mol Genet Metab. 2012. PMID: 22959828
-
ABCA1(Alabama): a novel variant associated with HDL deficiency and premature coronary artery disease.Atherosclerosis. 2002 Oct;164(2):245-50. doi: 10.1016/s0021-9150(02)00106-5. Atherosclerosis. 2002. PMID: 12204794 Review.
-
ABCA1-deficient mice: insights into the role of monocyte lipid efflux in HDL formation and inflammation.Arterioscler Thromb Vasc Biol. 2003 Jun 1;23(6):972-80. doi: 10.1161/01.ATV.0000054661.21499.FB. Epub 2003 Jan 9. Arterioscler Thromb Vasc Biol. 2003. PMID: 12615679 Review.
Cited by
-
Targeted inactivation of hepatic Abca1 causes profound hypoalphalipoproteinemia and kidney hypercatabolism of apoA-I.J Clin Invest. 2005 May;115(5):1333-42. doi: 10.1172/JCI23915. Epub 2005 Apr 7. J Clin Invest. 2005. PMID: 15841208 Free PMC article.
-
Ligand-dependent contribution of RXRbeta to cholesterol homeostasis in Sertoli cells.EMBO Rep. 2004 Mar;5(3):285-90. doi: 10.1038/sj.embor.7400094. EMBO Rep. 2004. PMID: 14993927 Free PMC article.
-
The ABCA subfamily--gene and protein structures, functions and associated hereditary diseases.Pflugers Arch. 2007 Feb;453(5):581-9. doi: 10.1007/s00424-006-0047-8. Epub 2006 Apr 4. Pflugers Arch. 2007. PMID: 16586097 Review.
-
Gene expression profiles of HTR8-S/Vneo cells after changes in ABCA1 expression.Funct Integr Genomics. 2018 Nov;18(6):725-735. doi: 10.1007/s10142-018-0621-0. Epub 2018 Jun 21. Funct Integr Genomics. 2018. PMID: 29931611
-
Transplacental Nutrient Transport Mechanisms of Intrauterine Growth Restriction in Rodent Models and Humans.Front Physiol. 2017 Nov 27;8:951. doi: 10.3389/fphys.2017.00951. eCollection 2017. Front Physiol. 2017. PMID: 29230179 Free PMC article. Review.
References
-
- Higgins CF: ABC-transporters: from microorganisms to man. Annu Rev Cell Biol 1992, 8:67-113 - PubMed
-
- Luciani MF, Denizot F, Savar S, Mattei MG, Chimini G: Cloning of two novel ABC transporters mapping on human chromosome 9. Genomics 1994, 21:150-159 - PubMed
-
- Connors TD, Van Raay TJ, Petry LR, Klinger KW, Landes GM, Burn TC: The cloning of a human ABC gene (ABC3) mapping to chromosome 16p13.3. Genomics 1997, 39:231–234 - PubMed
-
- Allikmets R, Singh N, Sun H, Shroyer NF, Hutchinson A, Chidambaram A, Gerrard B, Baird L, Stauffer D, Peiffer A, Rattner A, Smallwood P, Li Y, Anderson KL, Lewis RA, Nathans J, Leppert M, Dean M, Lupski JR: A photoreceptor cell-specific ATP-binding transporter gene (ABCR) is mutated in recessive Stargardt macular dystrophy. Nat Genet 1997, 15:236-246 - PubMed
-
- Riordan JR, Rommens JM, Kerem B, Alon N, Rozmahel R, Grzelczak Z, Zielenski J, Lok S, Plavsic N, Chou JL: Identification of the cystic fibrosis gene—cloning and characterization of complementary DNA. Science 1989, 245:1066-1072 - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
