

Association of human immunodeficiency virus type 1 Vif with RNA and its role in reverse transcription
- PMID: 10982337
- PMCID: PMC102089
- DOI: 10.1128/jvi.74.19.8938-8945.2000
Association of human immunodeficiency virus type 1 Vif with RNA and its role in reverse transcription
Abstract
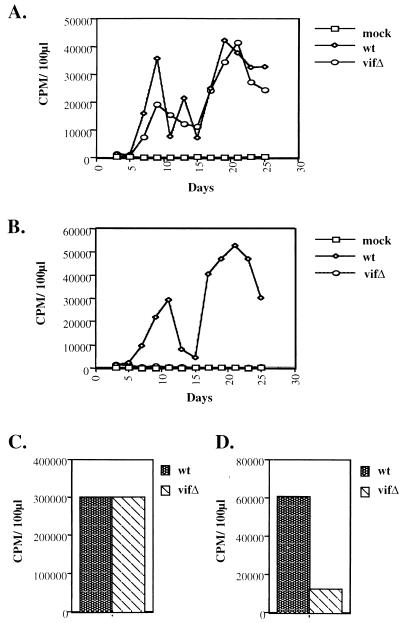
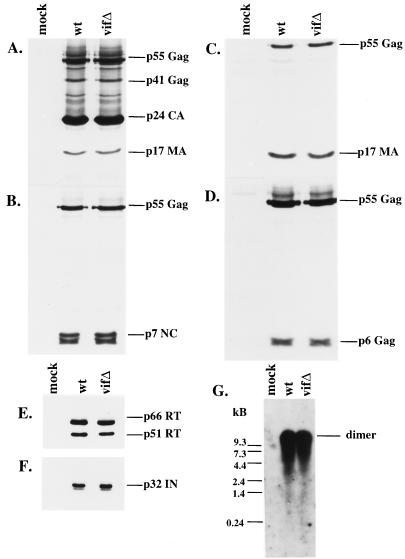
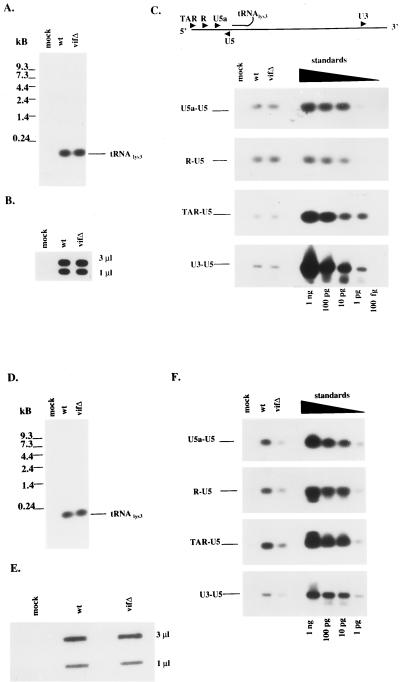
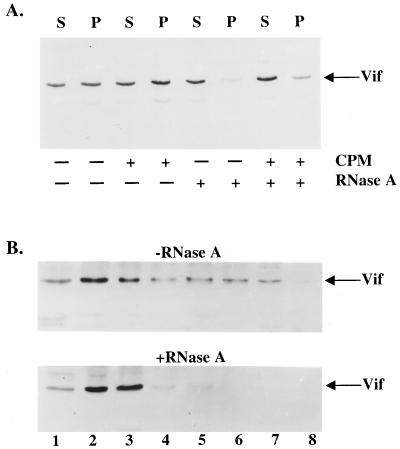
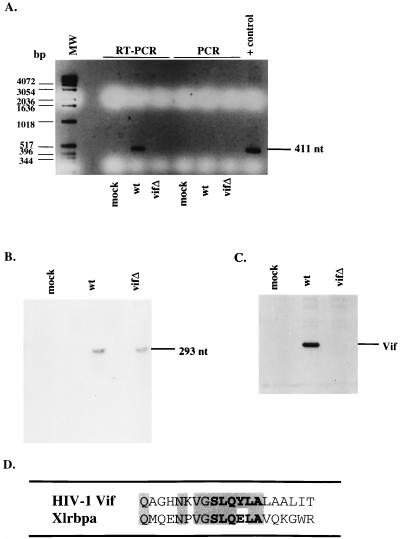
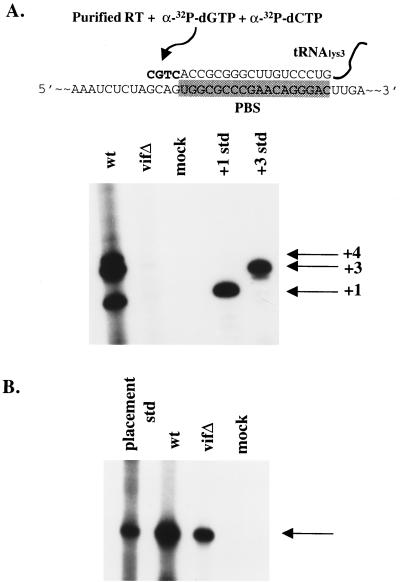
The vif gene of human immunodeficiency virus type 1 (HIV-1) is essential for viral replication, although the functional target of Vif remains elusive. HIV-1 vif mutant virions derived from nonpermissive H9 cells displayed no significant differences in the amount, ratio, or integrity of their protein composition relative to an isogenic wild-type virion. The amounts of the virion-associated viral genomic RNA and tRNA(3)(Lys) were additionally present at normal levels in vif mutant virions. We demonstrate that Vif associates with RNA in vitro as well as with viral genomic RNA in virus-infected cells. A functionally conserved lentivirus Vif motif was found in the double-stranded RNA binding domain of Xenopus laevis, Xlrbpa. The natural intravirion reverse transcriptase products were markedly reduced in vif mutant virions. Moreover, purified vif mutant genomic RNA-primer tRNA complexes displayed severe defects in the initiation of reverse transcription with recombinant reverse transcriptase. These data point to a novel role for Vif in the regulation of efficient reverse transcription through modulation of the virion nucleic acid components.
Figures
References
-
- Burd C G, Dreyfuss G. Conserved structures and diversity of functions of RNA-binding proteins. Science. 1994;265:615–621. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
