

Structure and activity of ClpB from Escherichia coli. Role of the amino-and -carboxyl-terminal domains
- PMID: 10982797
- PMCID: PMC1819559
- DOI: 10.1074/jbc.M005211200
Structure and activity of ClpB from Escherichia coli. Role of the amino-and -carboxyl-terminal domains
Abstract
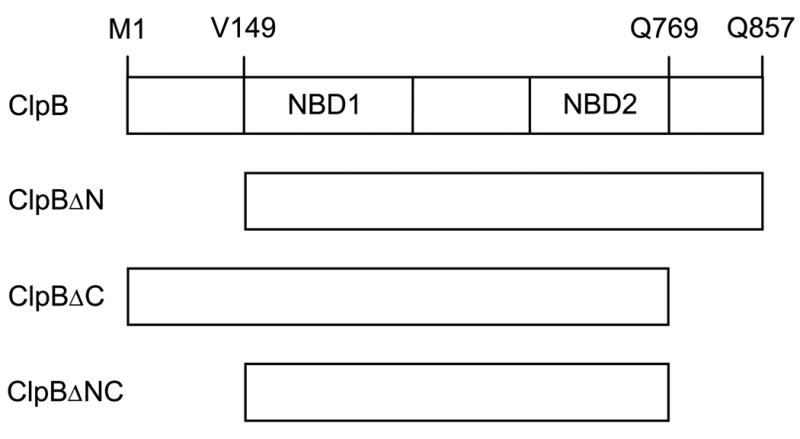
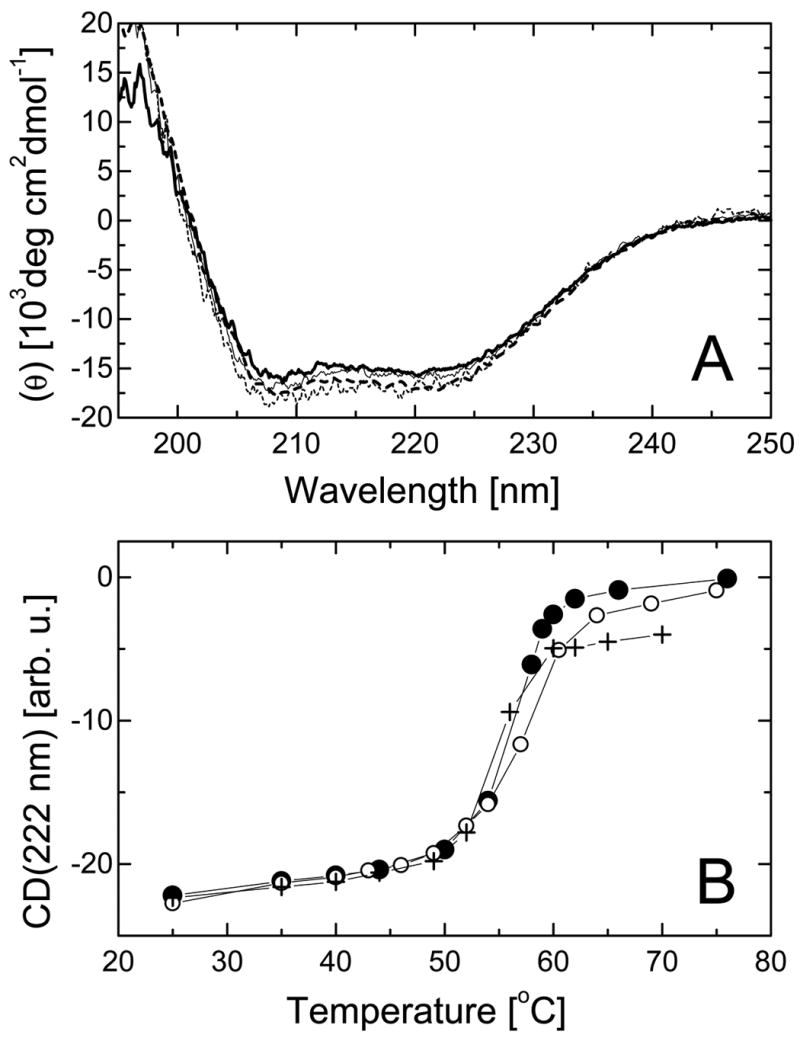
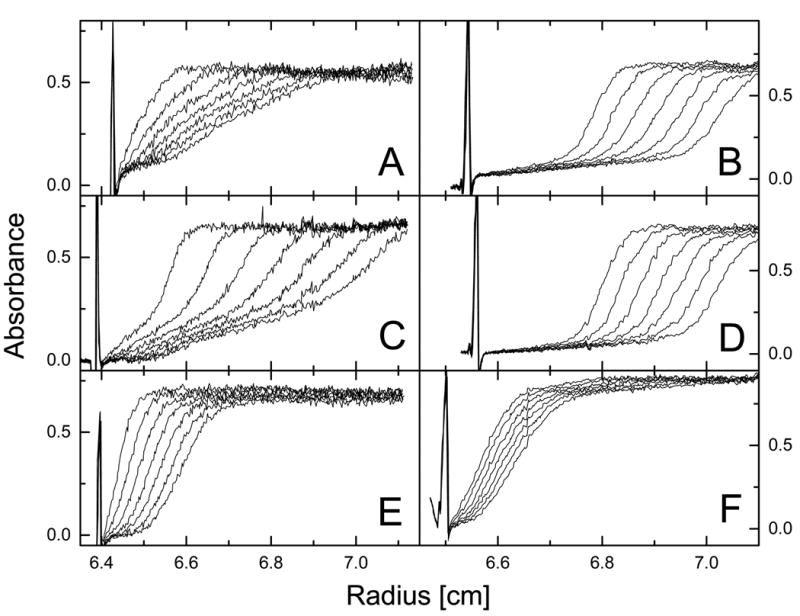
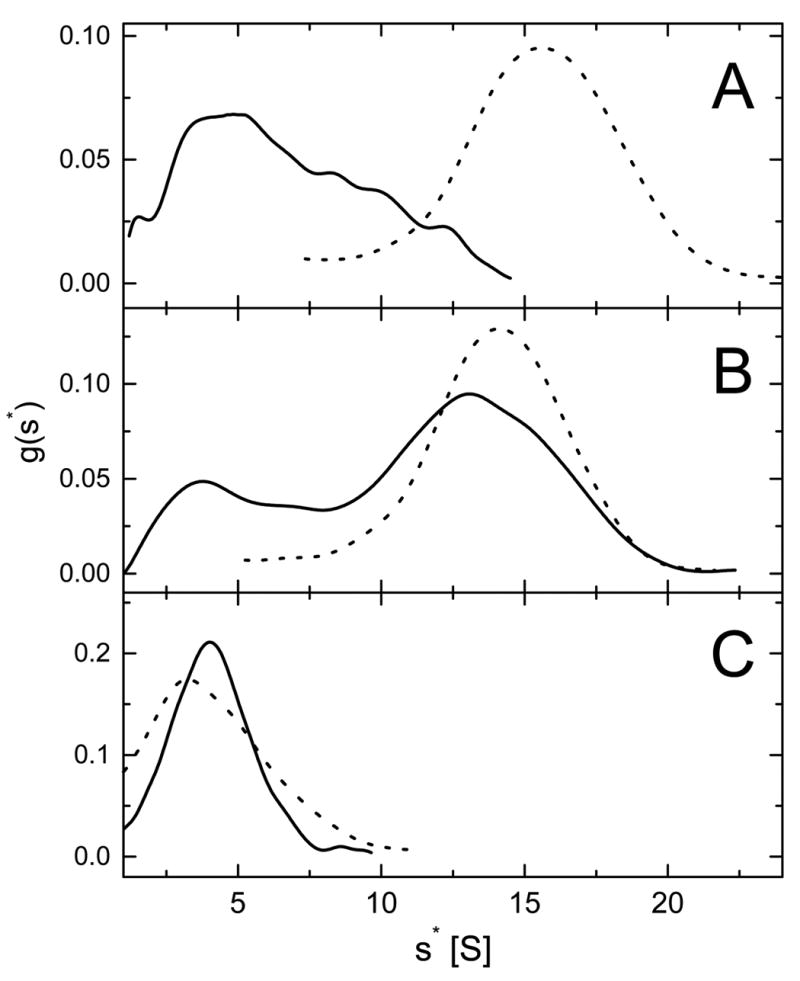
ClpB is a member of a protein-disaggregating multi-chaperone system in Escherichia coli. The mechanism of protein-folding reactions mediated by ClpB is currently unknown, and the functional role of different sequence regions in ClpB is under discussion. We have expressed and purified the full-length ClpB and three truncated variants with the N-terminal, C-terminal, and a double N- and C-terminal deletion. We studied the protein concentration-dependent and ATP-induced oligomerization of ClpB, casein-induced activation of ClpB ATPase, and ClpB-assisted reactivation of denatured firefly luciferase. We found that both the N- and C-terminal truncation of ClpB strongly inhibited its chaperone activity. The reasons for such inhibition were different, however, for the N- and C-terminal truncation. Deletion of the C-terminal domain inhibited the self-association of ClpB, which led to decreased affinity for ATP and to decreased ATPase and chaperone activity of the C-terminally truncated variants. In contrast, deletion of the N-terminal domain did not inhibit the self-association of ClpB and its basal ATPase activity but decreased the ability of casein to activate ClpB ATPase. These results indicate that the N-terminal region of ClpB may contain a functionally significant protein-binding site, whereas the main role of the C-terminal region is to support oligomerization of ClpB.
Figures
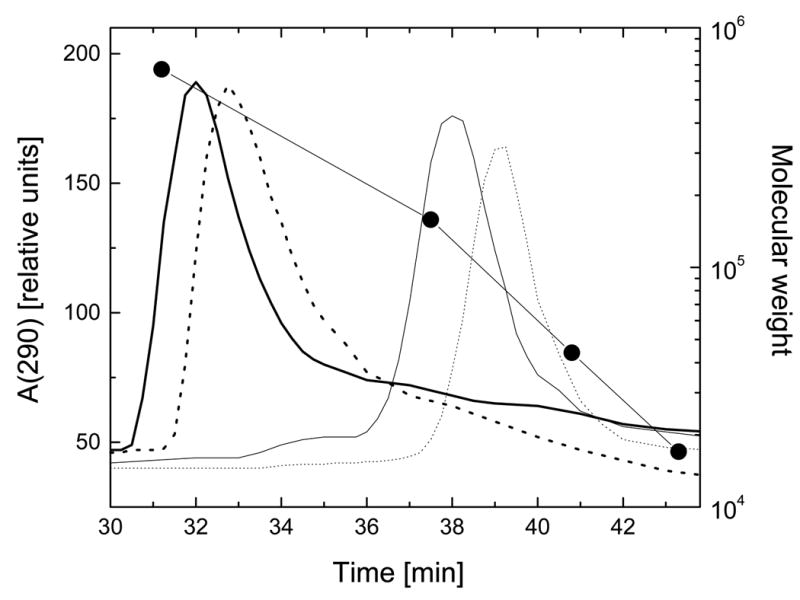
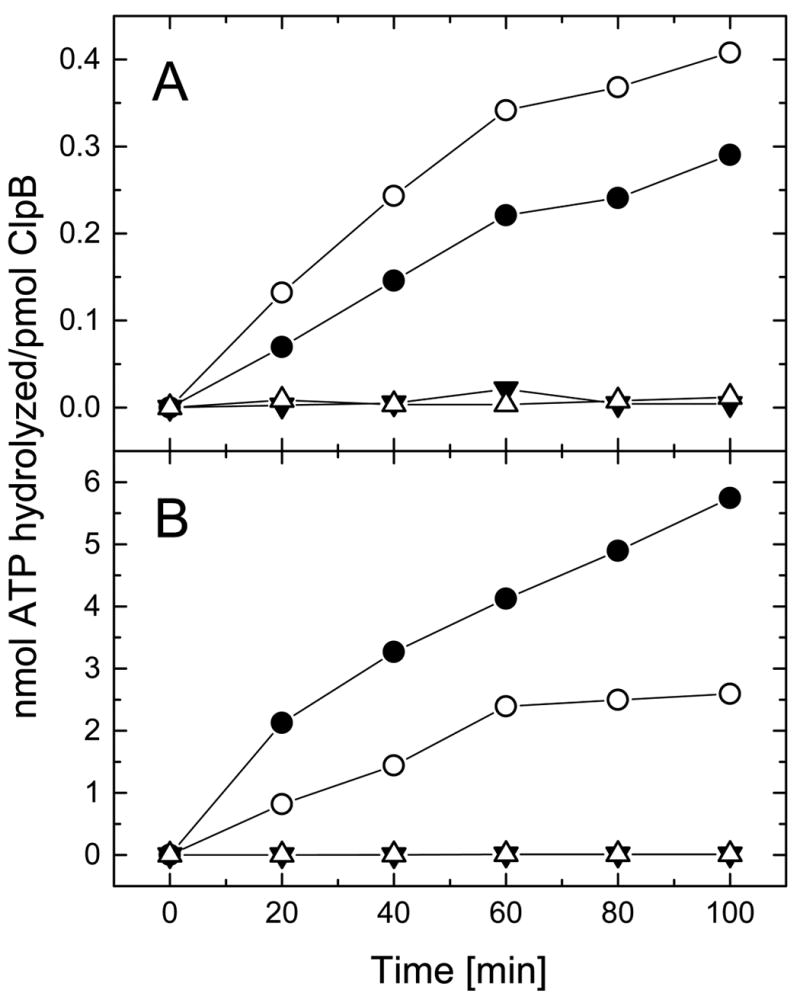

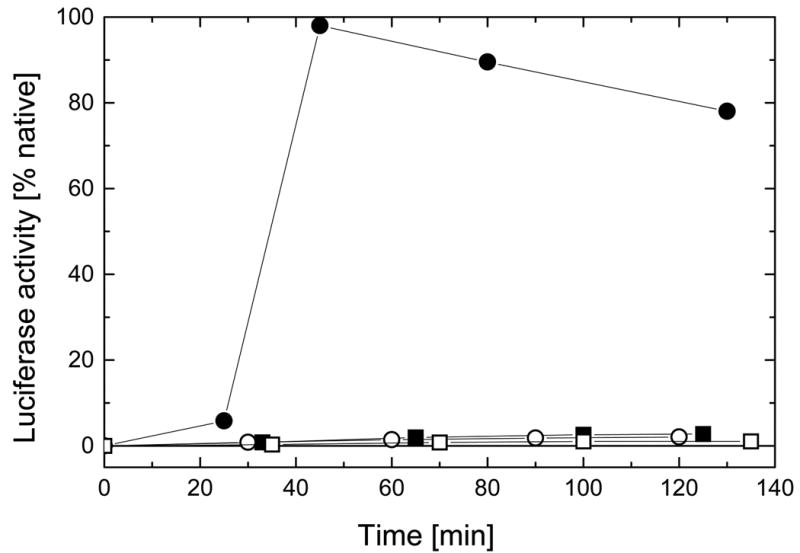
References
-
- Wickner S, Maurizi MR, Gottesman S. Science. 1999;286:1888–1893. - PubMed
-
- Schirmer EC, Glover JR, Singer MA, Lindquist S. Trends Biochem Sci. 1996;21:289–296. - PubMed
-
- Weber-Ban EU, Reid BG, Miranker AD, Horwich AL. Nature. 1999;401:90–93. - PubMed
-
- Zolkiewski M. J Biol Chem. 1999;274:28083–28086. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
