

Physical and functional interaction between the eukaryotic orthologs of prokaryotic translation initiation factors IF1 and IF2
- PMID: 10982835
- PMCID: PMC86272
- DOI: 10.1128/MCB.20.19.7183-7191.2000
Physical and functional interaction between the eukaryotic orthologs of prokaryotic translation initiation factors IF1 and IF2
Abstract
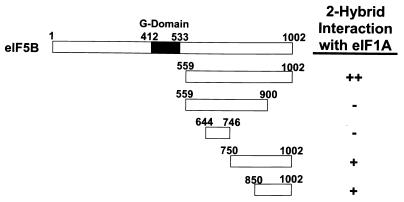
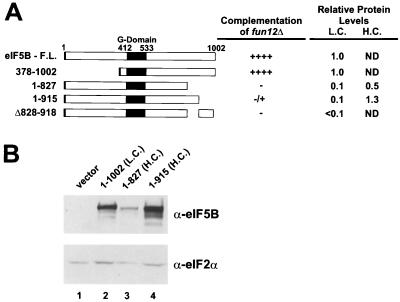
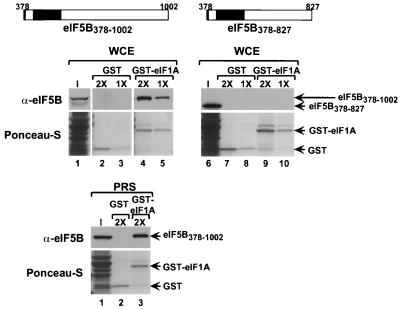
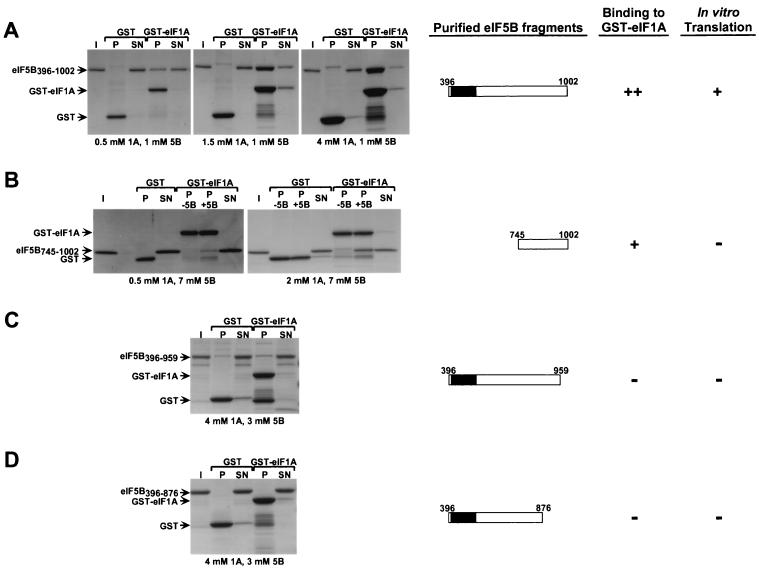
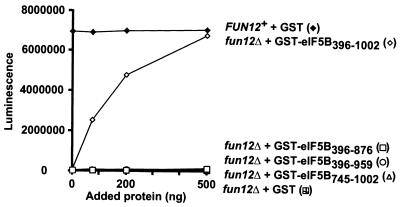
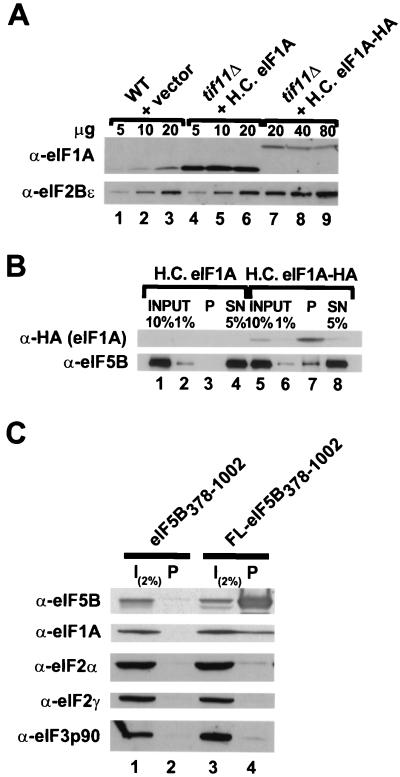
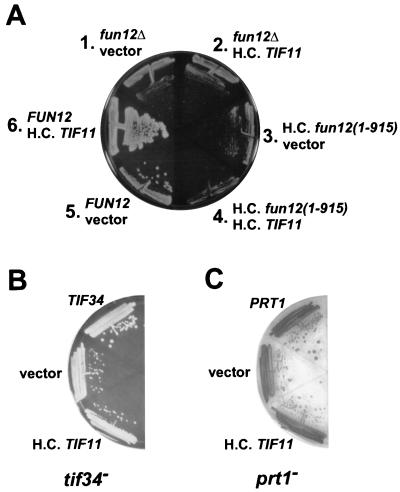
To initiate protein synthesis, a ribosome with bound initiator methionyl-tRNA must be assembled at the start codon of an mRNA. This process requires the coordinated activities of three translation initiation factors (IF) in prokaryotes and at least 12 translation initiation factors in eukaryotes (eIF). The factors eIF1A and eIF5B from eukaryotes show extensive amino acid sequence similarity to the factors IF1 and IF2 from prokaryotes. By a combination of two-hybrid, coimmunoprecipitation, and in vitro binding assays eIF1A and eIF5B were found to interact directly, and the eIF1A binding site was mapped to the C-terminal region of eIF5B. This portion of eIF5B was found to be critical for growth in vivo and for translation in vitro. Overexpression of eIF1A exacerbated the slow-growth phenotype of yeast strains expressing C-terminally truncated eIF5B. These findings indicate that the physical interaction between the evolutionarily conserved factors eIF1A and eIF5B plays an important role in translation initiation, perhaps to direct or stabilize the binding of methionyl-tRNA to the ribosomal P site.
Figures
Similar articles
-
Domains of eIF1A that mediate binding to eIF2, eIF3 and eIF5B and promote ternary complex recruitment in vivo.EMBO J. 2003 Jan 15;22(2):193-204. doi: 10.1093/emboj/cdg030. EMBO J. 2003. PMID: 12514125 Free PMC article.
-
Engaging the ribosome: universal IFs of translation.Trends Biochem Sci. 2001 Dec;26(12):705-9. doi: 10.1016/s0968-0004(01)02024-2. Trends Biochem Sci. 2001. PMID: 11738593
-
Mapping the binding interface between human eukaryotic initiation factors 1A and 5B: a new interaction between old partners.Proc Natl Acad Sci U S A. 2003 Feb 18;100(4):1535-40. doi: 10.1073/pnas.0437845100. Epub 2003 Feb 4. Proc Natl Acad Sci U S A. 2003. PMID: 12569173 Free PMC article.
-
Structure-function insights into prokaryotic and eukaryotic translation initiation.Curr Opin Struct Biol. 2009 Jun;19(3):300-9. doi: 10.1016/j.sbi.2009.04.010. Epub 2009 Jun 1. Curr Opin Struct Biol. 2009. PMID: 19493673 Review.
-
Molecular mechanisms of translation initiation in eukaryotes.Proc Natl Acad Sci U S A. 2001 Jun 19;98(13):7029-36. doi: 10.1073/pnas.111145798. Proc Natl Acad Sci U S A. 2001. PMID: 11416183 Free PMC article. Review.
Cited by
-
Domains of eIF1A that mediate binding to eIF2, eIF3 and eIF5B and promote ternary complex recruitment in vivo.EMBO J. 2003 Jan 15;22(2):193-204. doi: 10.1093/emboj/cdg030. EMBO J. 2003. PMID: 12514125 Free PMC article.
-
A single mammalian mitochondrial translation initiation factor functionally replaces two bacterial factors.Mol Cell. 2008 Feb 1;29(2):180-90. doi: 10.1016/j.molcel.2007.11.021. Mol Cell. 2008. PMID: 18243113 Free PMC article.
-
Eukaryotic Initiation Factor 5B (eIF5B) Cooperates with eIF1A and eIF5 to Facilitate uORF2-Mediated Repression of ATF4 Translation.Int J Mol Sci. 2018 Dec 13;19(12):4032. doi: 10.3390/ijms19124032. Int J Mol Sci. 2018. PMID: 30551605 Free PMC article.
-
Structural integrity of {alpha}-helix H12 in translation initiation factor eIF5B is critical for 80S complex stability.RNA. 2011 Apr;17(4):687-96. doi: 10.1261/rna.2412511. Epub 2011 Feb 18. RNA. 2011. PMID: 21335519 Free PMC article.
-
Translation initiation factor IF1 of Bacillus stearothermophilus and Thermus thermophilus substitute for Escherichia coli IF1 in vivo and in vitro without a direct IF1-IF2 interaction.Mol Microbiol. 2008 Dec;70(6):1368-77. doi: 10.1111/j.1365-2958.2008.06466.x. Epub 2008 Oct 2. Mol Microbiol. 2008. PMID: 18976282 Free PMC article.
References
-
- Asano K, Phan L, Anderson J, Hinnebusch A G. Complex formation by all five homologues of mammalian translation initiation factor 3 subunits from yeast Saccharomyces cerevisiae. J Biol Chem. 1998;273:18573–18585. - PubMed
-
- Battiste J B, Pestova T V, Hellen C U T, Wagner G. The eIF1A solution structure reveals a large RNA-binding surface important for scanning function. Mol Cell. 2000;5:109–119. - PubMed
-
- Benne R, Hershey J W B. The mechanism of action of protein synthesis initiation factors from rabbit reticulocytes. J Biol Chem. 1978;253:3078–3087. - PubMed
-
- Benne R, Naaktgeboren N, Gubbens J, Voorma H O. Recycling of initiation factors IF-1, IF-2 and IF-3. Eur J Biochem. 1973;32:372–380. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous