

Control of hnRNP A1 alternative splicing: an intron element represses use of the common 3' splice site
- PMID: 10982852
- PMCID: PMC86289
- DOI: 10.1128/MCB.20.19.7353-7362.2000
Control of hnRNP A1 alternative splicing: an intron element represses use of the common 3' splice site
Abstract
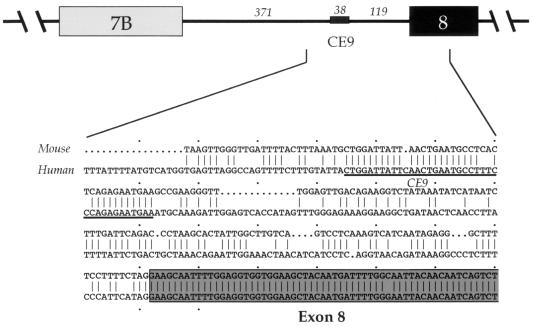
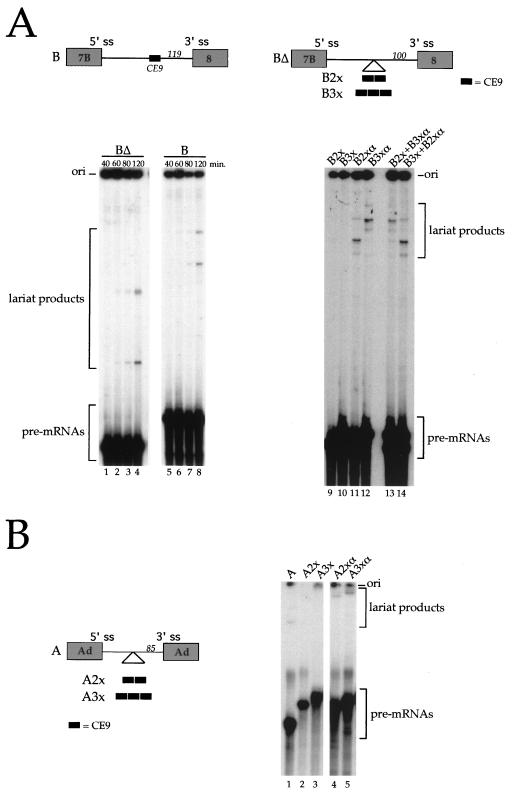
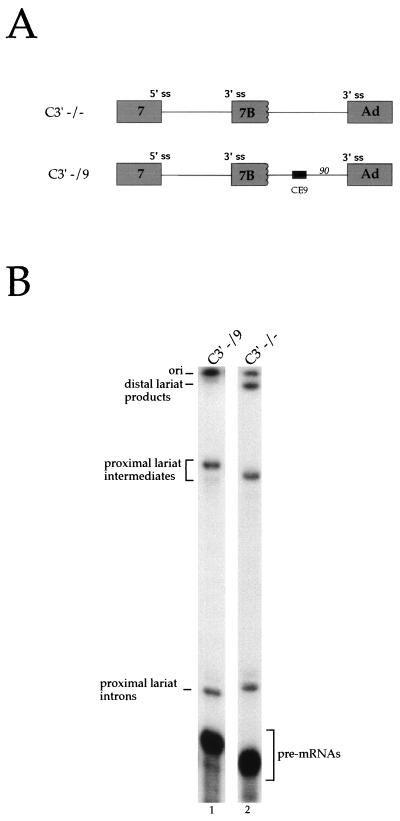
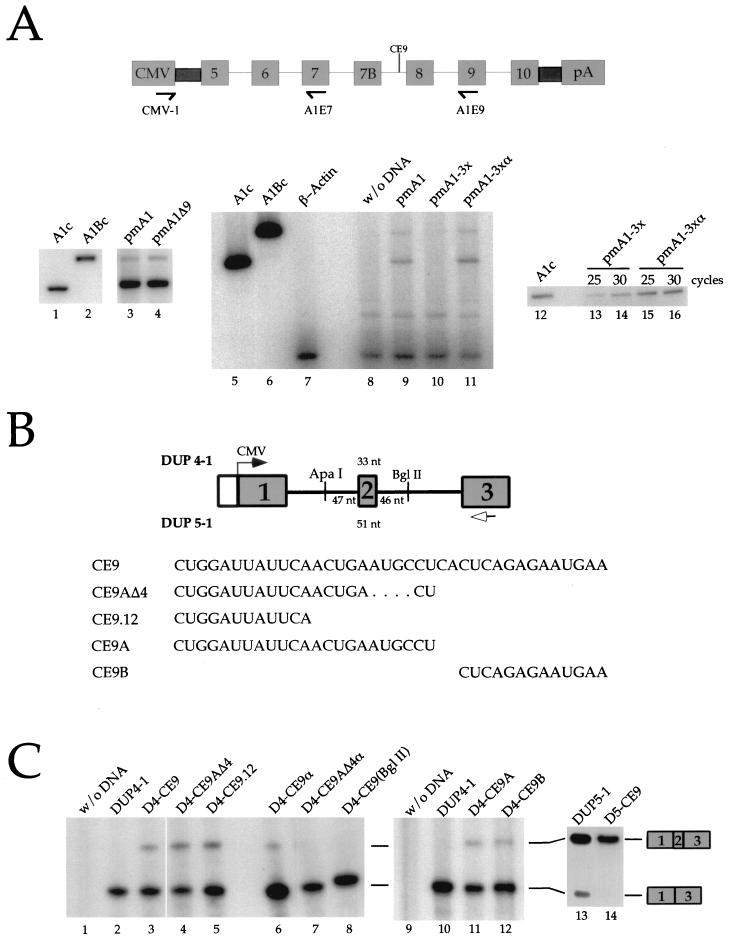
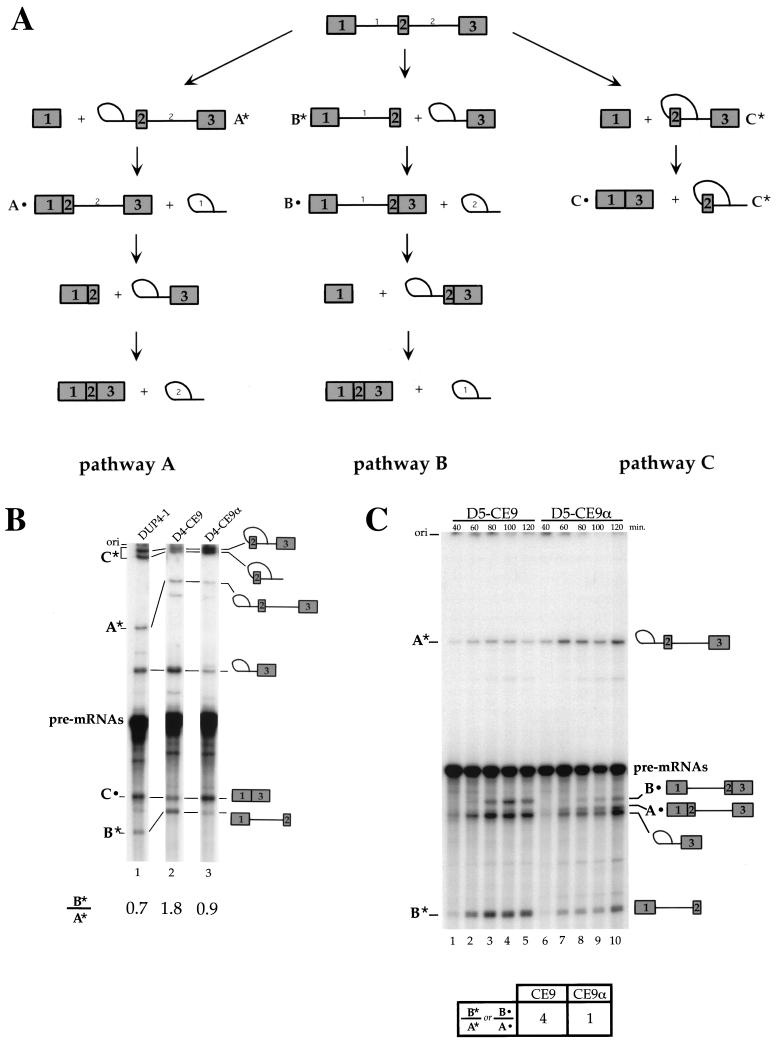
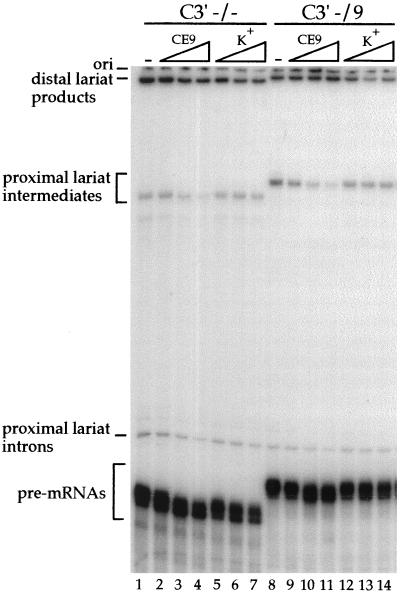
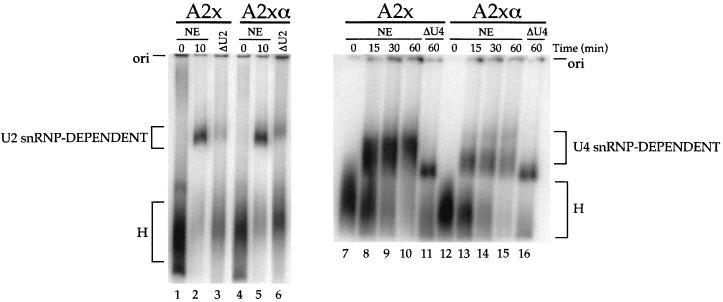
Alternative splicing of exon 7B in the hnRNP A1 pre-mRNA produces mRNAs encoding two proteins: hnRNP A1 and the less abundant A1B. We have reported the identification of several intron elements that contribute to exon 7B skipping. In this study, we report the activity of a novel element, conserved element 9 (CE9), located in the intron downstream of exon 7B. We show that multiple copies of CE9 inhibit exon 7B-exon 8 splicing in vitro. When CE9 is inserted between two competing 3' splice sites, a single copy of CE9 decreases splicing to the distal 3' splice site. Our in vivo results also support the conclusion that CE9 is a splicing modulator. First, inserting multiple copies of CE9 into an A1 minigene compromises the production of fully spliced products. Second, one copy of CE9 stimulates the inclusion of a short internal exon in a derivative of the human beta-globin gene. In this case, in vitro splicing assays suggest that CE9 decreases splicing of intron 1, an event that improves splicing of intron 2 and decreases skipping of the short internal exon. The ability of CE9 to act on heterologous substrates, combined with the results of a competition assay, suggest that the activity of CE9 is mediated by a trans-acting factor. Our results indicate that CE9 represses the use of the common 3' splice site in the hnRNP A1 alternative splicing unit.
Figures
Similar articles
-
SRp30c is a repressor of 3' splice site utilization.Mol Cell Biol. 2002 Jun;22(12):4001-10. doi: 10.1128/MCB.22.12.4001-4010.2002. Mol Cell Biol. 2002. PMID: 12024014 Free PMC article.
-
An intron element modulating 5' splice site selection in the hnRNP A1 pre-mRNA interacts with hnRNP A1.Mol Cell Biol. 1997 Apr;17(4):1776-86. doi: 10.1128/MCB.17.4.1776. Mol Cell Biol. 1997. PMID: 9121425 Free PMC article.
-
A highly stable duplex structure sequesters the 5' splice site region of hnRNP A1 alternative exon 7B.RNA. 1997 Apr;3(4):405-19. RNA. 1997. PMID: 9085847 Free PMC article.
-
Alternative splicing: multiple control mechanisms and involvement in human disease.Trends Genet. 2002 Apr;18(4):186-93. doi: 10.1016/s0168-9525(01)02626-9. Trends Genet. 2002. PMID: 11932019 Review.
-
hnRNP proteins and splicing control.Adv Exp Med Biol. 2007;623:123-47. doi: 10.1007/978-0-387-77374-2_8. Adv Exp Med Biol. 2007. PMID: 18380344 Review.
Cited by
-
Nanodiamonds coupled with 5,7-dimethoxycoumarin, a plant bioactive metabolite, interfere with the mitotic process in B16F10 cells altering the actin organization.Int J Nanomedicine. 2016 Feb 3;11:557-74. doi: 10.2147/IJN.S96614. eCollection 2016. Int J Nanomedicine. 2016. PMID: 26893562 Free PMC article.
-
SRp30c is a repressor of 3' splice site utilization.Mol Cell Biol. 2002 Jun;22(12):4001-10. doi: 10.1128/MCB.22.12.4001-4010.2002. Mol Cell Biol. 2002. PMID: 12024014 Free PMC article.
-
Oregano Phytocomplex Induces Programmed Cell Death in Melanoma Lines via Mitochondria and DNA Damage.Foods. 2020 Oct 17;9(10):1486. doi: 10.3390/foods9101486. Foods. 2020. PMID: 33080917 Free PMC article.
-
Redirecting splicing with bifunctional oligonucleotides.Nucleic Acids Res. 2014 Apr;42(6):e40. doi: 10.1093/nar/gkt1287. Epub 2013 Dec 26. Nucleic Acids Res. 2014. PMID: 24375754 Free PMC article.
-
hnRNP I/PTB can antagonize the splicing repressor activity of SRp30c.RNA. 2007 Aug;13(8):1287-300. doi: 10.1261/rna.403607. Epub 2007 Jun 4. RNA. 2007. PMID: 17548433 Free PMC article.
References
-
- Black D L. Activation of c-src neuron-specific splicing by an unusual RNA element in vivo and in vitro. Cell. 1992;69:795–807. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources