

Mutations in a tRNA import signal define distinct receptors at the two membranes of Leishmania mitochondria
- PMID: 10982857
- PMCID: PMC86294
- DOI: 10.1128/MCB.20.19.7410-7417.2000
Mutations in a tRNA import signal define distinct receptors at the two membranes of Leishmania mitochondria
Abstract
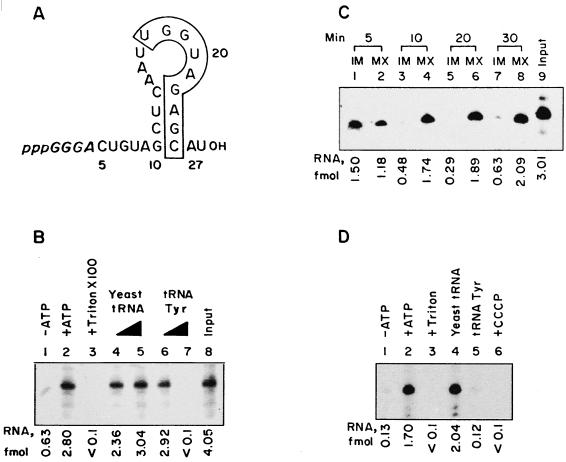
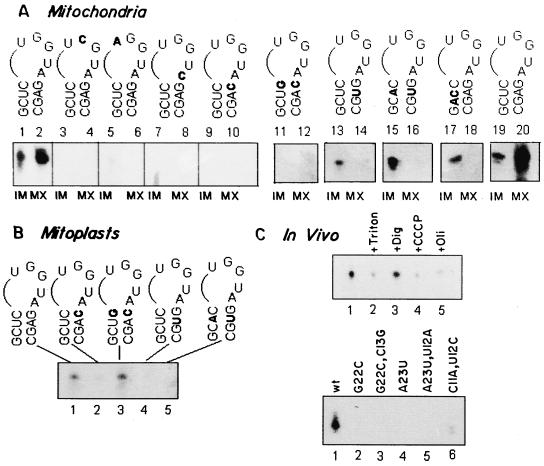
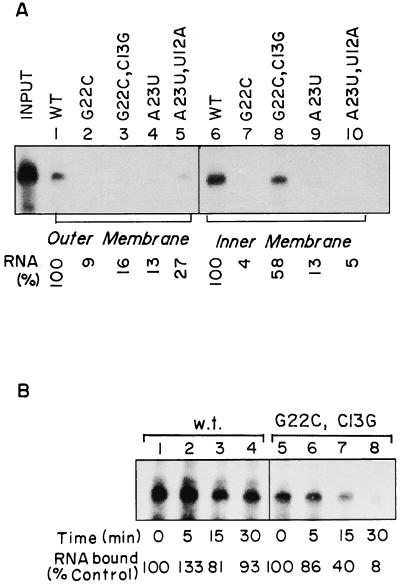
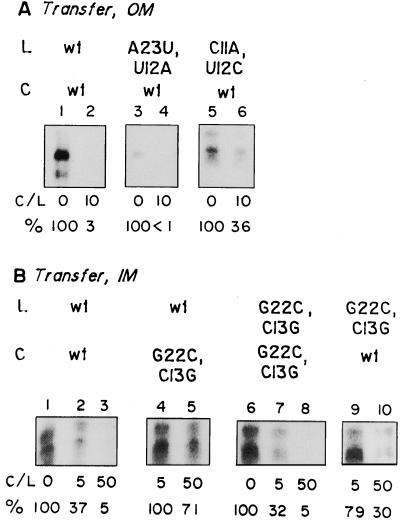
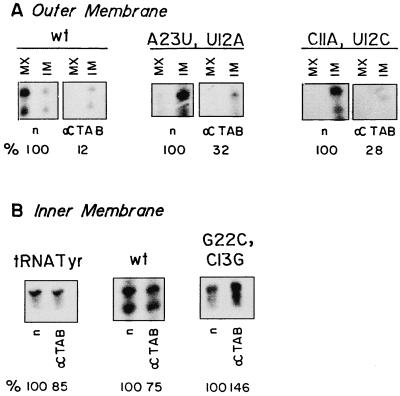
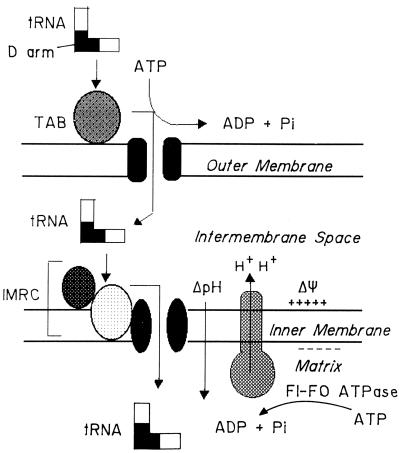
Nucleus-encoded tRNAs are selectively imported into the mitochondrion of Leishmania, a kinetoplastid protozoan. An oligoribonucleotide constituting the D stem-loop import signal of tRNA(Tyr)(GUA) was efficiently transported into the mitochondrial matrix in organello as well as in vivo. Transfer through the inner membrane could be uncoupled from that through the outer membrane and was resistant to antibody against the outer membrane receptor TAB. A number of mutations in the import signal had differential effects on outer and inner membrane transfer. Some mutants which efficiently traversed the outer membrane were unable to enter the matrix. Conversely, restoration of the loop-closing GC pair in reverse resulted in reversion of transfer through the inner, but not the outer, membrane, and binding of the RNA to the inner membrane was restored. These experiments indicate the presence at the two membranes of receptors with distinct specificities which mediate stepwise transfer into the mitochondrial matrix. The combination of oligonucleotide mutagenesis and biochemical fractionation may provide a general tool for the identification of tRNA transport factors.
Figures
Similar articles
-
Mitochondrial RNA import in Leishmania tropica: aptamers homologous to multiple tRNA domains that interact cooperatively or antagonistically at the inner membrane.Mol Cell Biol. 2002 Jun;22(12):4372-82. doi: 10.1128/MCB.22.12.4372-4382.2002. Mol Cell Biol. 2002. PMID: 12024047 Free PMC article.
-
Stepwise transfer of tRNA through the double membrane of Leishmania mitochondria.J Biol Chem. 1999 Oct 29;274(44):31249-55. doi: 10.1074/jbc.274.44.31249. J Biol Chem. 1999. PMID: 10531321
-
Role of an RNA-binding protein in import of tRNA into Leishmania mitochondria.J Biol Chem. 1997 Aug 22;272(34):21396-402. doi: 10.1074/jbc.272.34.21396. J Biol Chem. 1997. PMID: 9261154
-
Leishmania mitochondrial tRNA importers.Int J Biochem Cell Biol. 2008;40(12):2681-5. doi: 10.1016/j.biocel.2007.10.025. Epub 2007 Nov 20. Int J Biochem Cell Biol. 2008. PMID: 18061510 Review.
-
Import and insertion of proteins into the mitochondrial outer membrane.Eur J Biochem. 1995 Jan 15;227(1-2):9-18. doi: 10.1111/j.1432-1033.1995.tb20354.x. Eur J Biochem. 1995. PMID: 7851447 Review.
Cited by
-
"Ping-pong" interactions between mitochondrial tRNA import receptors within a multiprotein complex.Mol Cell Biol. 2003 Aug;23(15):5217-24. doi: 10.1128/MCB.23.15.5217-5224.2003. Mol Cell Biol. 2003. PMID: 12861008 Free PMC article.
-
Necessary and sufficient factors for the import of transfer RNA into the kinetoplast mitochondrion.EMBO Rep. 2007 Jun;8(6):589-95. doi: 10.1038/sj.embor.7400979. Epub 2007 May 18. EMBO Rep. 2007. PMID: 17510656 Free PMC article.
-
Proton-guided movements of tRNA within the Leishmania mitochondrial RNA import complex.Nucleic Acids Res. 2008 Mar;36(5):1599-609. doi: 10.1093/nar/gkn010. Epub 2008 Feb 3. Nucleic Acids Res. 2008. PMID: 18250088 Free PMC article.
-
A bifunctional tRNA import receptor from Leishmania mitochondria.Proc Natl Acad Sci U S A. 2006 May 30;103(22):8354-9. doi: 10.1073/pnas.0510869103. Epub 2006 May 19. Proc Natl Acad Sci U S A. 2006. Retraction in: Proc Natl Acad Sci U S A. 2025 Jul 29;122(30):e2517057122. doi: 10.1073/pnas.2517057122. PMID: 16714384 Free PMC article. Retracted.
-
Mitochondrial RNA import in Leishmania tropica: aptamers homologous to multiple tRNA domains that interact cooperatively or antagonistically at the inner membrane.Mol Cell Biol. 2002 Jun;22(12):4372-82. doi: 10.1128/MCB.22.12.4372-4382.2002. Mol Cell Biol. 2002. PMID: 12024047 Free PMC article.
References
-
- Adhya S, Ghosh T, Das A, Bera S K, Mahapatra S. Role of an RNA binding protein in import of tRNA into Leishmania mitochondria. J Biol Chem. 1997;272:21396–21402. - PubMed
-
- Chiu N, Chiu A, Suyama Y. Native and imported transfer RNA in mitochondria. J Mol Biol. 1975;94:37–50. - PubMed
-
- Das S, Adhya S. Organization and chromosomal localization of β-tubulin genes in Leishmania donovani. J Biosci. 1990;15:239–248.
-
- Gray M W, Burger G, Lang B F. Mitochondrial evolution. Science. 1999;283:1476–1481. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous