

Molecular basis for CD40 signaling mediated by TRAF3
- PMID: 10984535
- PMCID: PMC27035
- DOI: 10.1073/pnas.97.19.10395
Molecular basis for CD40 signaling mediated by TRAF3
Abstract
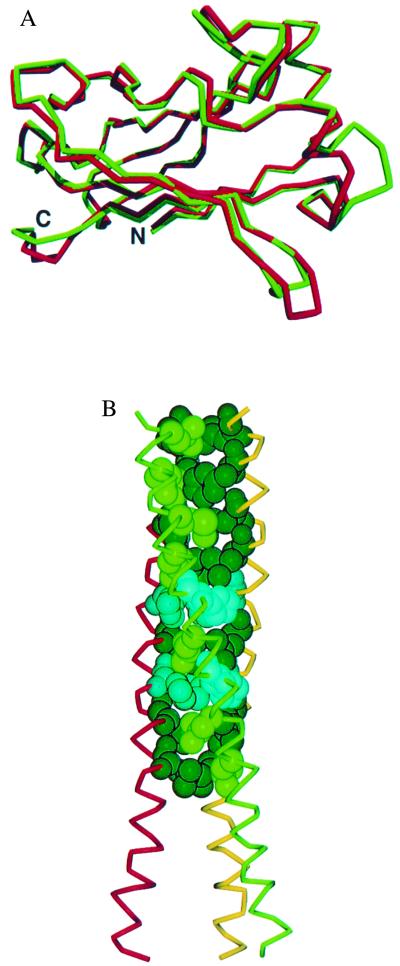
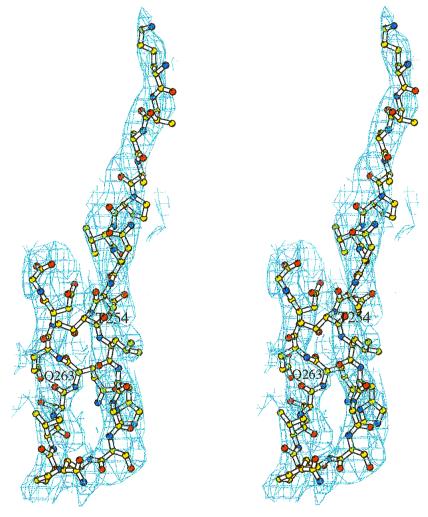
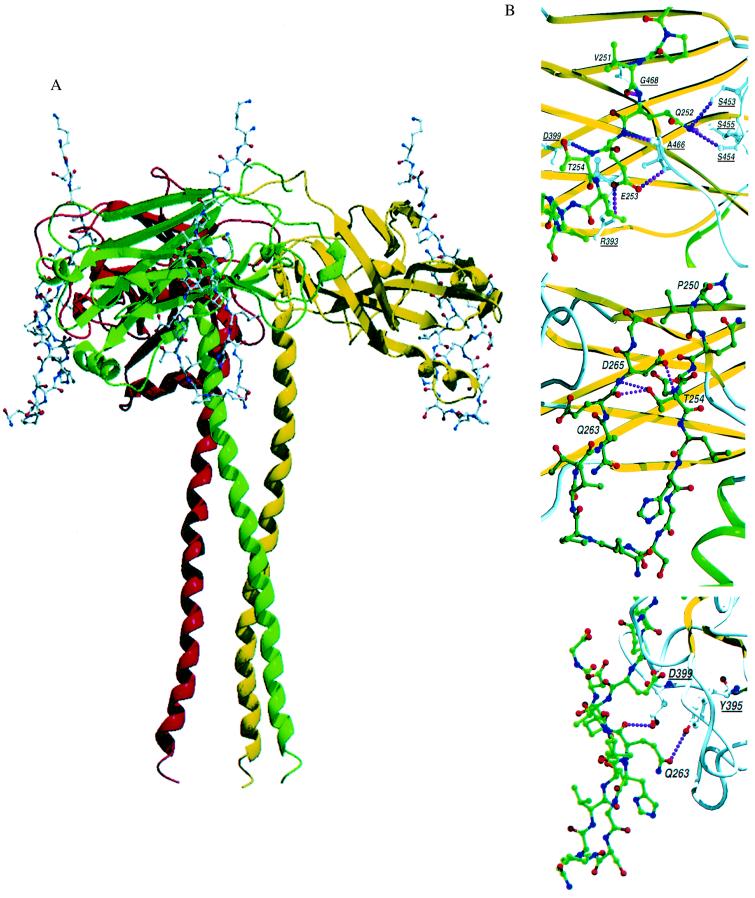
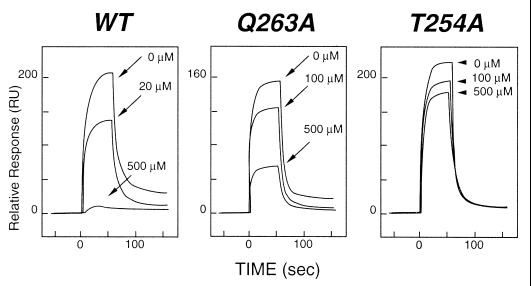
Tumor necrosis factor receptors (TNFR) are single transmembrane-spanning glycoproteins that bind cytokines and trigger multiple signal transduction pathways. Many of these TNFRs rely on interactions with TRAF proteins that bind to the intracellular domain of the receptors. CD40 is a member of the TNFR family that binds to several different TRAF proteins. We have determined the crystal structure of a 20-residue fragment from the cytoplasmic domain of CD40 in complex with the TRAF domain of TRAF3. The CD40 fragment binds as a hairpin loop across the surface of the TRAF domain. Residues shown by mutagenesis and deletion analysis to be critical for TRAF3 binding are involved either in direct contact with TRAF3 or in intramolecular interactions that stabilize the hairpin. Comparison of the interactions of CD40 with TRAF3 vs. TRAF2 suggests that CD40 may assume different conformations when bound to different TRAF family members. This molecular adaptation may influence binding affinity and specific cellular triggers.
Figures
References
-
- Foy T M, Aruffo A, Bajorath J, Buhlmann J E, Noelle R J. Annu Rev Immunol. 1996;14:591–617. - PubMed
-
- Banchereau J, Bazan F, Blanchard D, Briere F, Galizzi J P, van Kooten C, Liu Y J, Rousset F, Saeland S. Annu Rev Immunol. 1994;12:881–922. - PubMed
-
- Clark E A, Ledbetter J A. Nature (London) 1994;367:425–428. - PubMed
-
- Eliopoulos A G, Dawson C W, Mosialos G, Floettmann J E, Rowe M, Armitage R J, Dawson J, Zapata J M, Kerr D J, Wakelam M J, et al. Oncogene. 1996;13:2243–2254. - PubMed
-
- Eliopoulos A G, Stack M, Dawson C W, Kaye K M, Hodgkin L, Sihota S, Rowe M, Young L S. Oncogene. 1997;14:2899–2916. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
