

Rapid generation of nested chromosomal deletions on mouse chromosome 2
- PMID: 10984539
- PMCID: PMC27048
- DOI: 10.1073/pnas.97.19.10471
Rapid generation of nested chromosomal deletions on mouse chromosome 2
Abstract
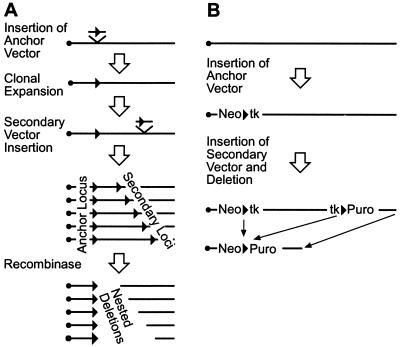
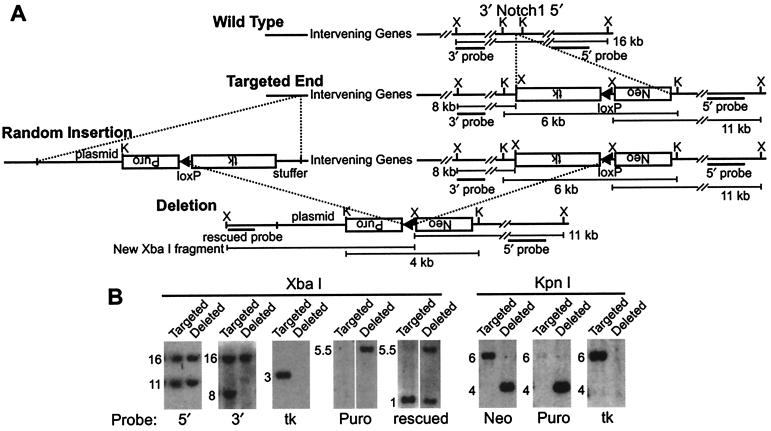
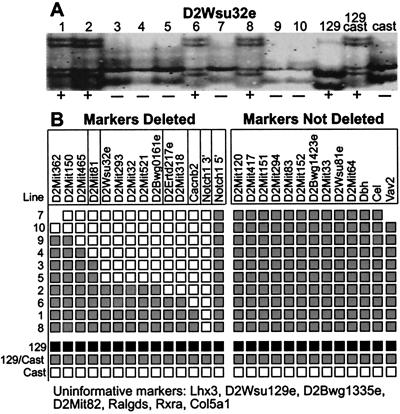
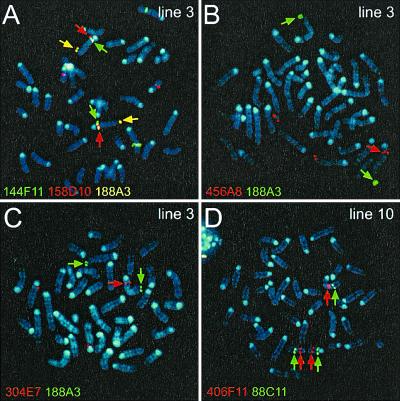
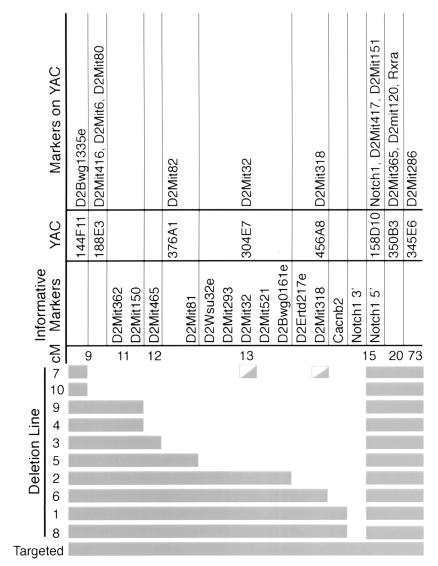
Nested chromosomal deletions are powerful genetic tools. They are particularly suited for identifying essential genes in development either directly or by screening induced mutations against a deletion. To apply this approach to the functional analysis of mouse chromosome 2, a strategy for the rapid generation of nested deletions with Cre recombinase was developed and tested. A loxP site was targeted to the Notch1 gene on chromosome 2. A targeted line was cotransfected with a second loxP site and a plasmid for transient expression of Cre. Independent random integrations of the second loxP site onto the targeted chromosome in direct repeat orientation created multiple nested deletions. By virtue of targeting in an F(1) hybrid embryonic stem cell line, F(1)(129S1xCast/Ei), the deletions could be verified and rapidly mapped. Ten deletions fell into seven size classes, with the largest extending six or seven centiMorgans. The cytology of the deletion chromosomes were determined by fluorescent in situ hybridization. Eight deletions were cytologically normal, but the two largest deletions had additional rearrangements. Three deletions, including the largest unrearranged deletion, have been transmitted through the germ line. Several endpoints also have been cloned by plasmid rescue. These experiments illustrate the means to rapidly create and map deletions anywhere in the mouse genome. They also demonstrate an improved method for generating nested deletions in embryonic stem cells.
Figures
Similar articles
-
Nested chromosomal deletions induced with retroviral vectors in mice.Nat Genet. 2000 Jan;24(1):92-5. doi: 10.1038/71756. Nat Genet. 2000. PMID: 10615136
-
X-ray-induced mutations in mouse embryonic stem cells.Proc Natl Acad Sci U S A. 1998 Feb 3;95(3):1114-9. doi: 10.1073/pnas.95.3.1114. Proc Natl Acad Sci U S A. 1998. PMID: 9448294 Free PMC article.
-
A site-directed chromosomal translocation induced in embryonic stem cells by Cre-loxP recombination.Nat Genet. 1995 Apr;9(4):376-85. doi: 10.1038/ng0495-376. Nat Genet. 1995. PMID: 7795643
-
Engineering chromosomal rearrangements in mice.Nat Rev Genet. 2001 Oct;2(10):780-90. doi: 10.1038/35093564. Nat Rev Genet. 2001. PMID: 11584294 Review.
-
From mouse to man: generating megabase chromosome rearrangements.Trends Genet. 2001 Jun;17(6):331-9. doi: 10.1016/s0168-9525(01)02321-6. Trends Genet. 2001. PMID: 11377795 Review.
Cited by
-
A p53-dependent mechanism underlies macrocytic anemia in a mouse model of human 5q- syndrome.Nat Med. 2010 Jan;16(1):59-66. doi: 10.1038/nm.2063. Epub 2009 Nov 22. Nat Med. 2010. PMID: 19966810 Free PMC article.
-
X-ray-induced deletion complexes in embryonic stem cells on mouse chromosome 15.Mamm Genome. 2005 Sep;16(9):661-71. doi: 10.1007/s00335-005-0011-5. Epub 2005 Oct 20. Mamm Genome. 2005. PMID: 16245023
-
Development of an enhanced GFP-based dual-color reporter to facilitate genetic screens for the recovery of mutations in mice.Proc Natl Acad Sci U S A. 2003 Nov 25;100(24):14103-8. doi: 10.1073/pnas.1936166100. Epub 2003 Nov 13. Proc Natl Acad Sci U S A. 2003. PMID: 14615591 Free PMC article.
-
Modeling chromosomes in mouse to explore the function of genes, genomic disorders, and chromosomal organization.PLoS Genet. 2006 Jul;2(7):e86. doi: 10.1371/journal.pgen.0020086. PLoS Genet. 2006. PMID: 16839184 Free PMC article. Review.
-
Effect of divergence time and recombination rate on molecular evolution of Drosophila INE-1 transposable elements and other candidates for neutrally evolving sites.J Mol Evol. 2007 Dec;65(6):627-39. doi: 10.1007/s00239-007-9028-6. Epub 2007 Sep 25. J Mol Evol. 2007. PMID: 17896069
References
-
- You Y, Browning V L, Schimenti J C. Methods. 1997;13:409–421. - PubMed
-
- Rinchik E, Russell L B. In: Genome Analysis. Davies K, Tilghman S, editors. Vol. 1. Plainview, NY: Cold Spring Harbor Lab. Press; 1990. pp. 121–158.
-
- Holdener-Kenny B, Sharan S, Magnuson T. BioEssays. 1992;14:831–838. - PubMed
-
- Justice M J, Zheng B, Woychik R P, Bradley A. Methods. 1997;13:423–436. - PubMed
-
- Russell L B. Mutat Res. 1971;11:107–123. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
