

Activation of the beta globin locus by transcription factors and chromatin modifiers
- PMID: 10990462
- PMCID: PMC314215
- DOI: 10.1093/emboj/19.18.4986
Activation of the beta globin locus by transcription factors and chromatin modifiers
Abstract
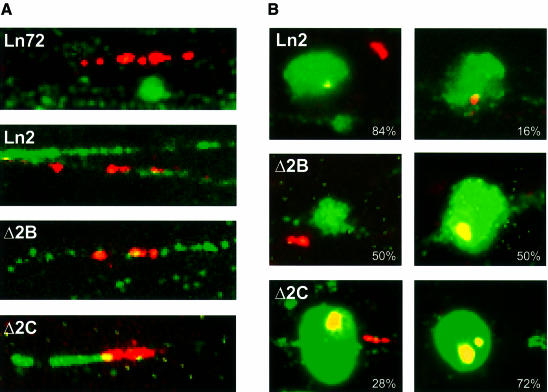
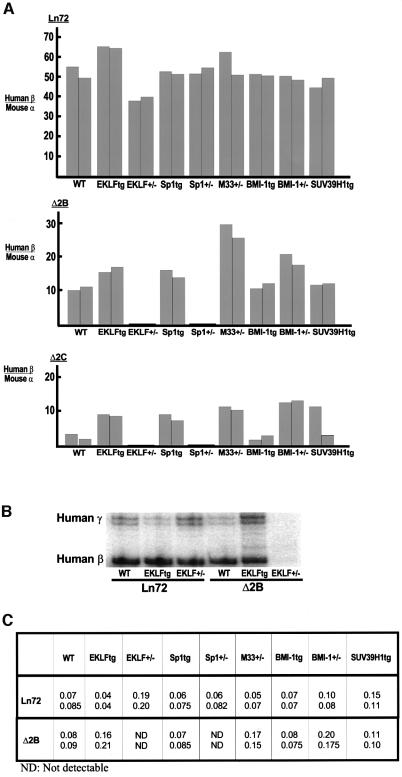
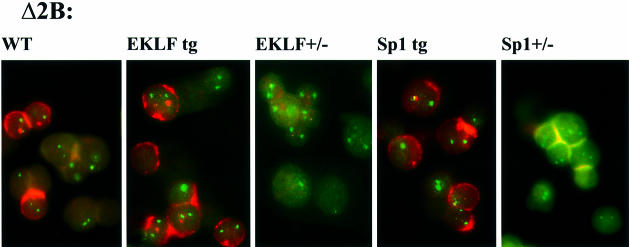
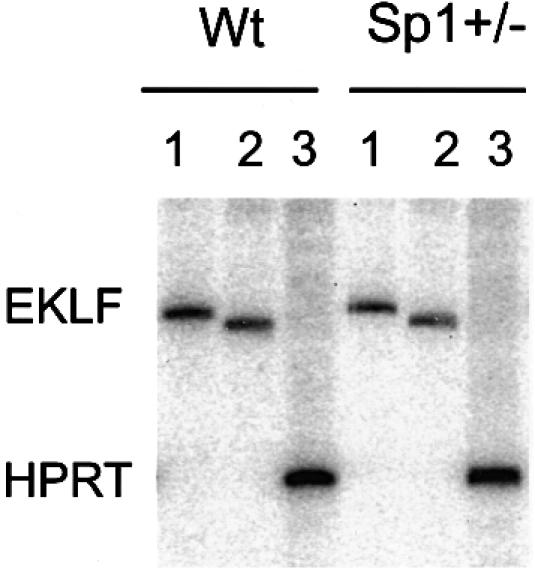
Locus control regions (LCRs) alleviate chromatin-mediated transcriptional repression. Incomplete LCRs partially lose this property when integrated in transcriptionally restrictive genomic regions such as centromeres. This frequently results in position effect variegation (PEV), i.e. the suppression of expression in a proportion of the cells. Here we show that this PEV is influenced by the heterochromatic protein SUV39H1 and by the Polycomb group proteins M33 and BMI-1. A concentration variation of these proteins modulates the proportion of cells expressing human globins in a locus-dependent manner. Similarly, the transcription factors Sp1 or erythroid Krüppel-like factor (EKLF) also influence PEV, characterized by a change in the number of expressing cells and the chromatin structure of the locus. However, in contrast to results obtained in a euchromatic locus, EKLF influences the expression of the gamma- more than the beta-globin genes, suggesting that the relief of silencing is caused by the binding of EKLF to the LCR and that genes at an LCR proximal position are more likely to be in an open chromatin state than genes at a distal position.
Figures
References
-
- Aagaard L., Schmid,M., Warburton,P. and Jenuwein,T. (2000) Mitotic phosphorylation of SUV39H1, a novel component of active centromeres, coincides with transient accumulation at mammalian centromeres. J. Cell Sci., 113, 817–829. - PubMed
-
- Alami R., Greally,J.M., Tanimoto,K., Hwang,S., Feng,Y.Q., Engel,J.D., Fiering,S. and Bouhassira,E.E. (2000) β-globin YAC transgenes exhibit uniform expression levels but position effect variegation in mice. Hum. Mol. Genet., 9, 631–636. - PubMed
-
- Alkema M.J., van der Lugt,N.M.T., Bobeldijk,R.C., Berns,A. and van Lohuizen,M. (1995) Transformation of axial skeleton due to overexpression of bmi-1 in transgenic mice. Nature, 374, 724–727. - PubMed
-
- Aparicio O.M. and Gottschling,D.E. (1994) Overcoming telomeric silencing: a trans-activator competes to establish gene expression in a cell cycle-dependent way. Genes Dev., 8, 1133–1146. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
