

Intrinsic constraint on plasmablast growth and extrinsic limits of plasma cell survival
- PMID: 10993912
- PMCID: PMC2193289
- DOI: 10.1084/jem.192.6.813
Intrinsic constraint on plasmablast growth and extrinsic limits of plasma cell survival
Abstract
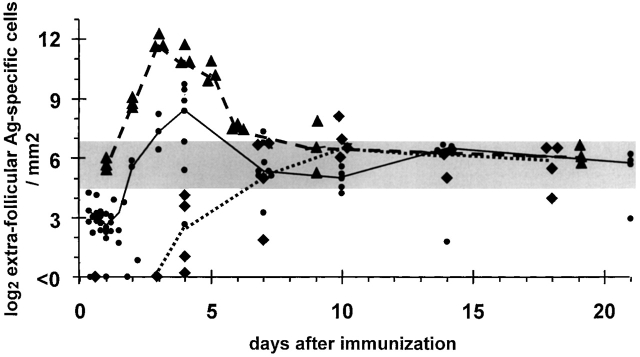
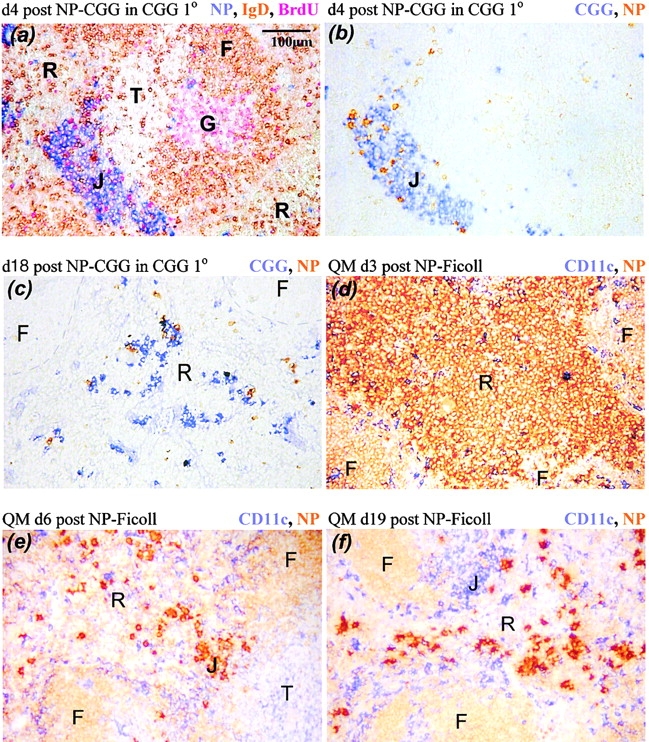
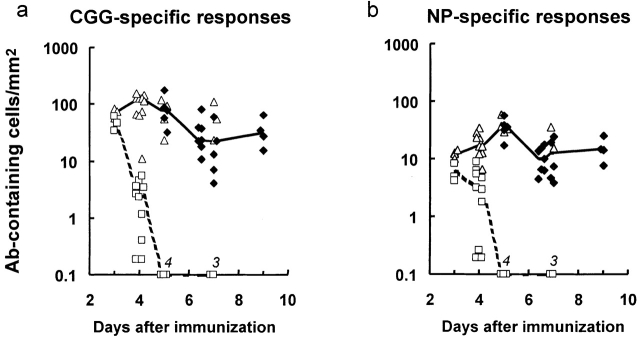
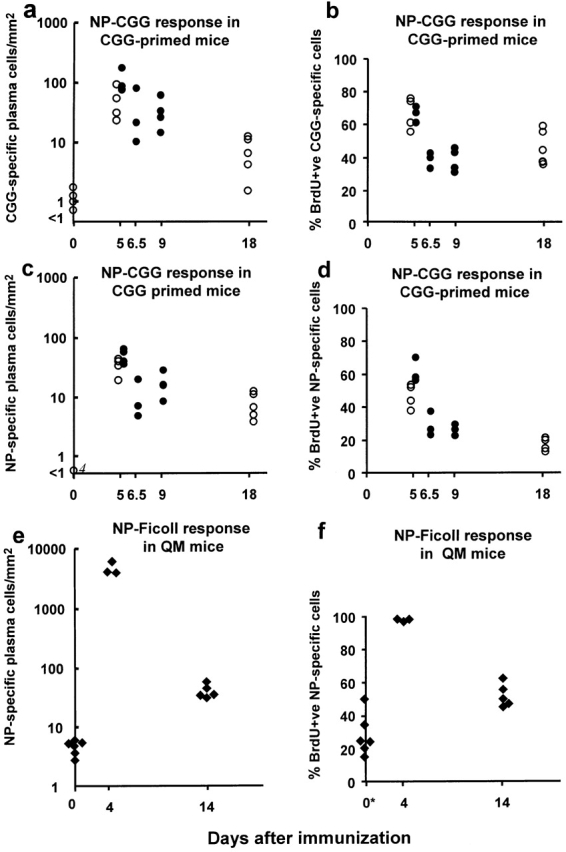
B cells recruited into splenic antibody responses grow exponentially, either in extrafollicular foci as plasmablasts, or in follicles where they form germinal centers. Both responses yield plasma cells. Although many splenic plasma cells survive <3 d, some live much longer. This study shows that early plasma cell death relates to a finite capacity of the spleen to sustain plasma cells rather than a life span endowed by the cell's origin or the quality of antibody it produces. Antibody responses were compared where the peak numbers of plasma cells in spleen sections varied between 100 and 5,000 cells/mm(2). In each response, plasmablast clones divided some five times, with the peak numbers of plasma cells produced relating directly to the number of B cells recruited into the response. The spleen seems to have the capacity to sustain between 20 and 100 plasma cells/mm(2). When this number is exceeded, there is a loss of excess cells. Immunoglobulin variable region gene sequencing, and 5-bromo-2'-deoxyuridine pulse-chase studies indicate that long-lived splenic plasma cells are a mixture of cells derived from the extrafollicular and germinal center responses and cells derived from virgin and memory B cells. Only a proportion has switched immunoglobulin class.
Figures


References
-
- Liu Y.J., Zhang J., Lane P.J.L., Chan E.Y.-T., MacLennan I.C.M. Sites of specific B cell activation in primary and secondary responses to T cell-dependent and T cell-independent antigens. Eur. J. Immunol. 1991;21:2951–2962. - PubMed
-
- García de Vinuesa C., Gulbranson-Judge A., Khan M., O'Leary P., Cascalho M., Wabl M., Klaus G.G.B., Owen M.J., MacLennan I.C.M. Dendritic cells associated with plasmablast survival. Eur. J. Immunol. 1999;29:3712–3721. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
