

HFR1 encodes an atypical bHLH protein that acts in phytochrome A signal transduction
- PMID: 10995393
- PMCID: PMC316929
HFR1 encodes an atypical bHLH protein that acts in phytochrome A signal transduction
Abstract
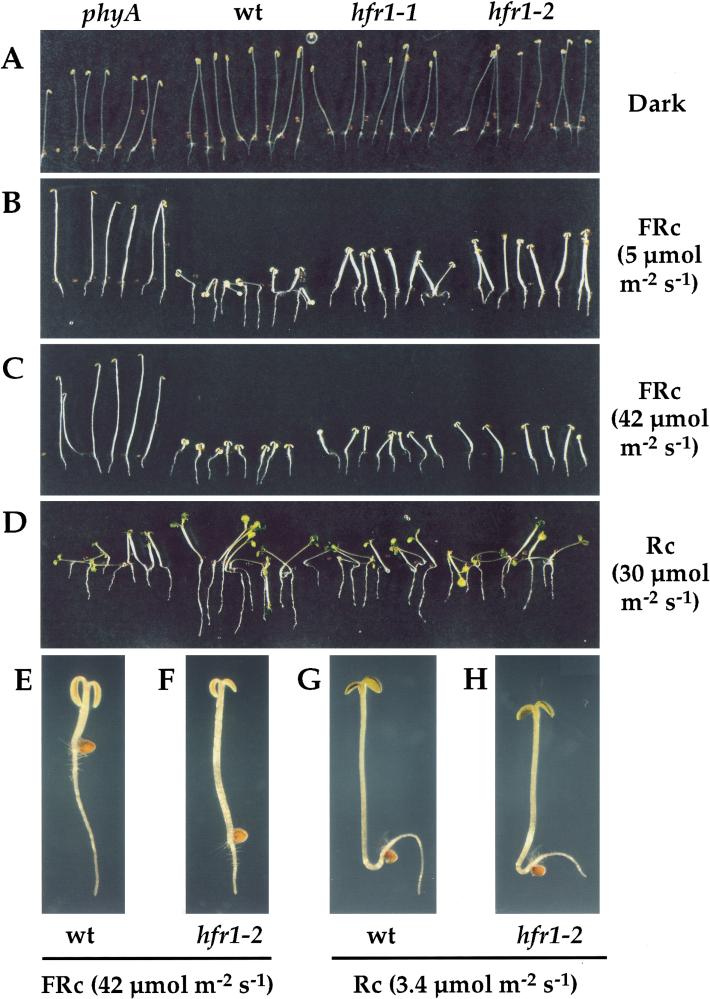
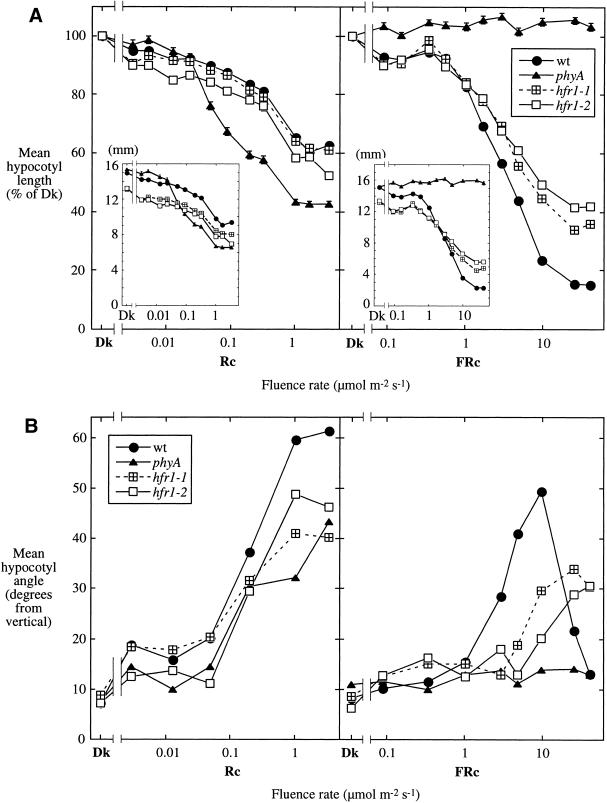
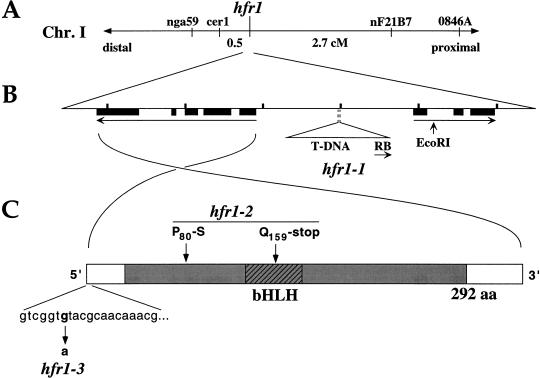
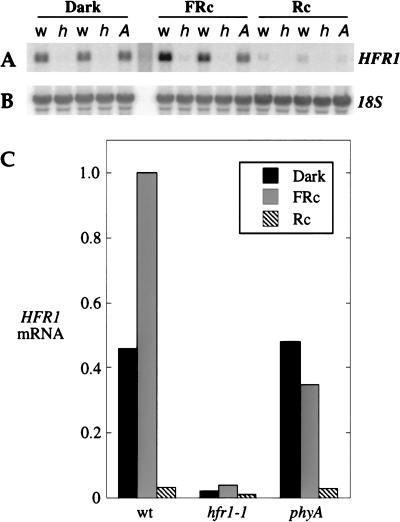
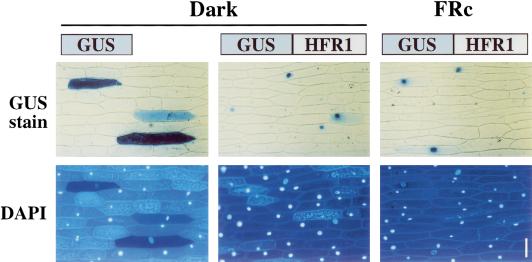
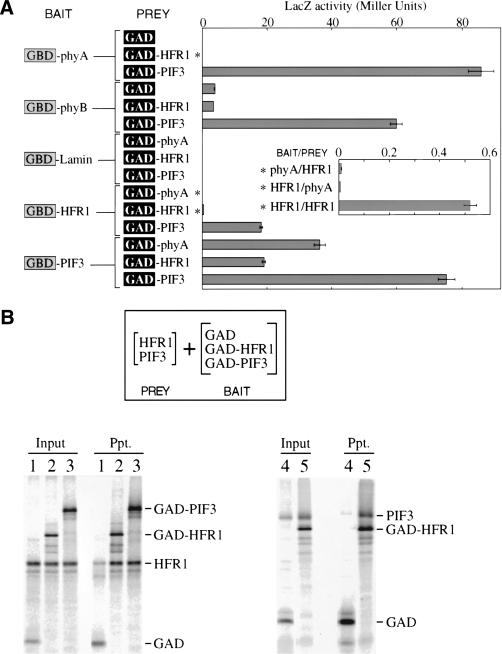
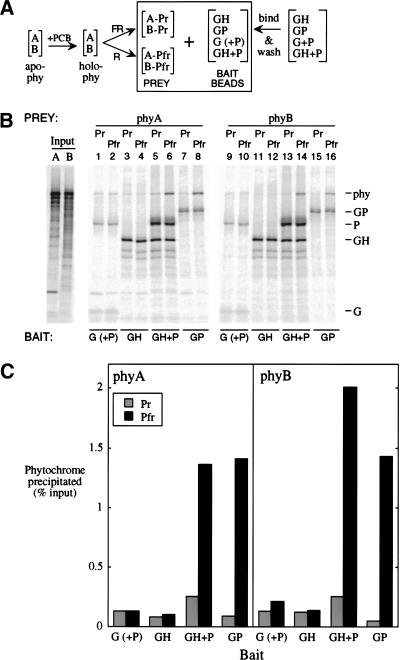
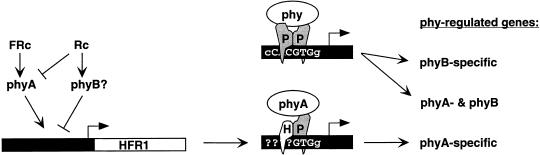
Phytochromes are informational photoreceptors through which plants adapt their growth and development to prevailing light conditions. These adaptations are effected primarily through phytochrome regulation of gene expression by mechanisms that remain unclear. We describe a new mutant, hfr1 (long hypocotyl in far-red), that exhibits a reduction in seedling responsiveness specifically to continuous far-red light (FRc), thereby suggesting a locus likely to be involved in phytochrome A (phyA) signal transduction. Using an insertionally tagged allele, we cloned the HFR1 gene and subsequently confirmed its identity with additional alleles derived from a directed genetic screen. HFR1 encodes a nuclear protein with strong similarity to the bHLH family of DNA-binding proteins but with an atypical basic region. In contrast to PIF3, a related bHLH protein previously shown to bind phyB, HFR1 did not bind either phyA or B. However, HFR1 did bind PIF3, suggesting heterodimerization, and both the HFR1/PIF3 complex and PIF3 homodimer bound preferentially to the Pfr form of both phytochromes. Thus, HFR1 may function to modulate phyA signaling via heterodimerization with PIF3. HFR1 mRNA is 30-fold more abundant in FRc than in continuous red light, suggesting a potential mechanistic basis for the specificity of HFR1 to phyA signaling.
Figures

References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases