

The reduction of chromosome number in meiosis is determined by properties built into the chromosomes
- PMID: 10995430
- PMCID: PMC2150703
- DOI: 10.1083/jcb.150.6.1223
The reduction of chromosome number in meiosis is determined by properties built into the chromosomes
Abstract
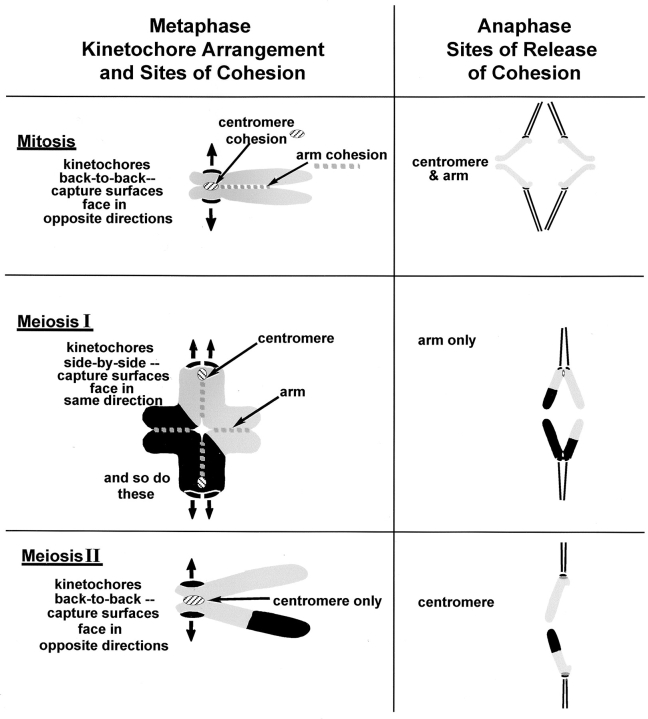
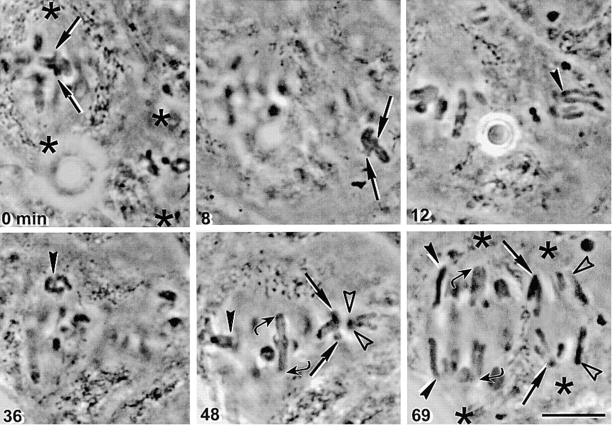
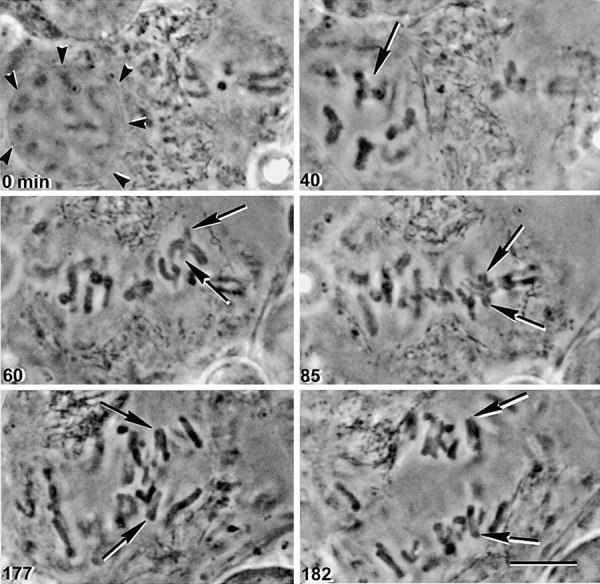
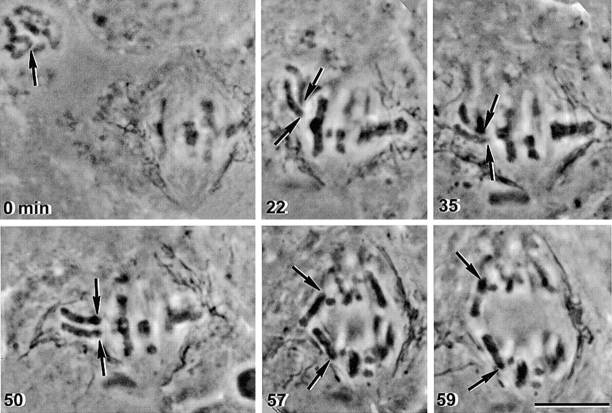
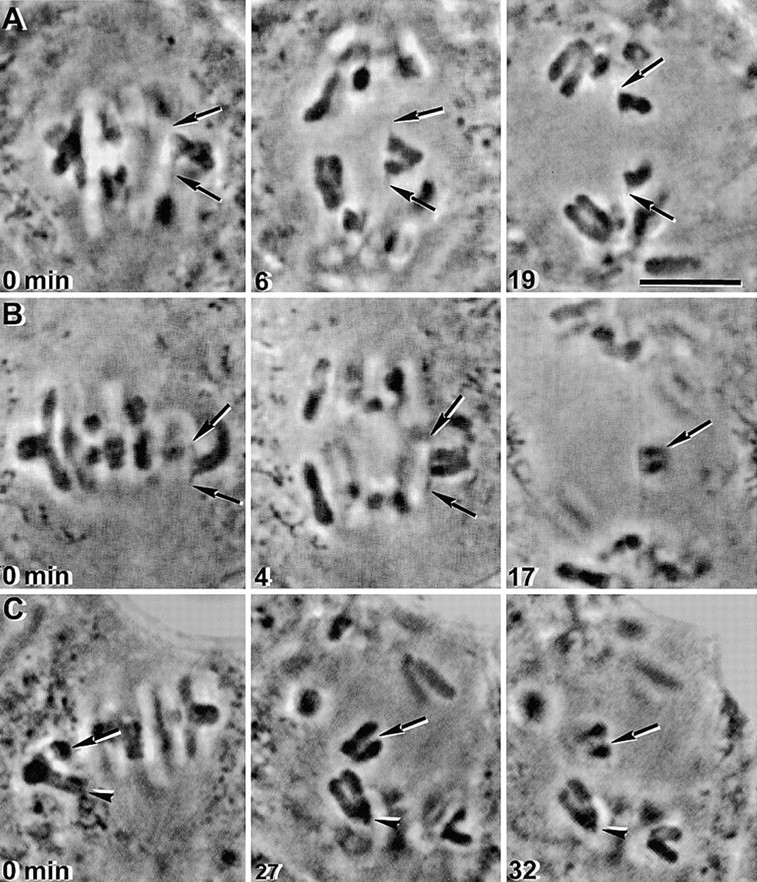
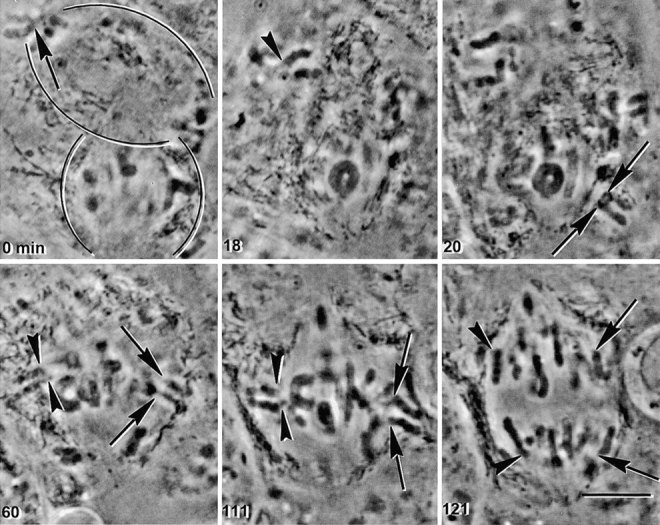
In meiosis I, two chromatids move to each spindle pole. Then, in meiosis II, the two are distributed, one to each future gamete. This requires that meiosis I chromosomes attach to the spindle differently than meiosis II chromosomes and that they regulate chromosome cohesion differently. We investigated whether the information that dictates the division type of the chromosome comes from the whole cell, the spindle, or the chromosome itself. Also, we determined when chromosomes can switch from meiosis I behavior to meiosis II behavior. We used a micromanipulation needle to fuse grasshopper spermatocytes in meiosis I to spermatocytes in meiosis II, and to move chromosomes from one spindle to the other. Chromosomes placed on spindles of a different meiotic division always behaved as they would have on their native spindle; e.g., a meiosis I chromosome attached to a meiosis II spindle in its normal fashion and sister chromatids moved together to the same spindle pole. We also showed that meiosis I chromosomes become competent meiosis II chromosomes in anaphase of meiosis I, but not before. The patterns for attachment to the spindle and regulation of cohesion are built into the chromosome itself. These results suggest that regulation of chromosome cohesion may be linked to differences in the arrangement of kinetochores in the two meiotic divisions.
Figures
References
-
- Bickel S.E., Orr-Weaver T.L. Holding chromatids together to ensure they go their separate ways. Bioessays. 1996;18:293–300. - PubMed
-
- Gimenez-Abian J.F., Clarke D.J., Garcia de la Vega C., Gimenez-Martin G. The role of sister chromatid cohesiveness and structure in meiotic behaviour. Chromosoma. 1997;106:422–434. - PubMed
-
- Goldstein L.S. Kinetochore structure and its role in chromosome orientation during the first meiotic division in male D. melanogaster . Cell. 1981;25:591–602. - PubMed
