

Desmin cytoskeleton linked to muscle mitochondrial distribution and respiratory function
- PMID: 10995435
- PMCID: PMC2150713
- DOI: 10.1083/jcb.150.6.1283
Desmin cytoskeleton linked to muscle mitochondrial distribution and respiratory function
Abstract
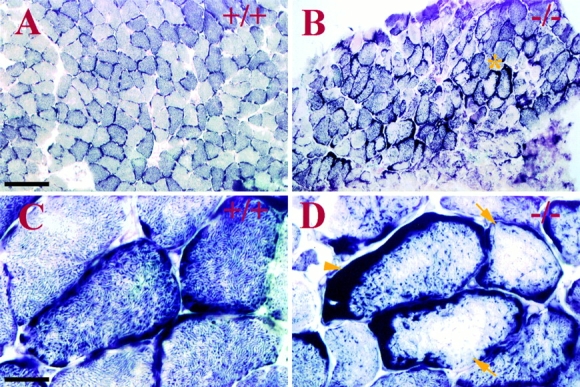
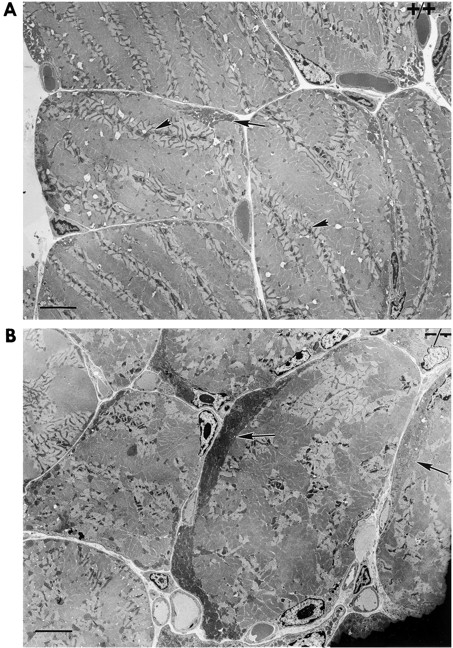
Ultrastructural studies have previously suggested potential association of intermediate filaments (IFs) with mitochondria. Thus, we have investigated mitochondrial distribution and function in muscle lacking the IF protein desmin. Immunostaining of skeletal muscle tissue sections, as well as histochemical staining for the mitochondrial marker enzymes cytochrome C oxidase and succinate dehydrogenase, demonstrate abnormal accumulation of subsarcolemmal clumps of mitochondria in predominantly slow twitch skeletal muscle of desmin-null mice. Ultrastructural observation of desmin-null cardiac muscle demonstrates in addition to clumping, extensive mitochondrial proliferation in a significant fraction of the myocytes, particularly after work overload. These alterations are frequently associated with swelling and degeneration of the mitochondrial matrix. Mitochondrial abnormalities can be detected very early, before other structural defects become obvious. To investigate related changes in mitochondrial function, we have analyzed ADP-stimulated respiration of isolated muscle mitochondria, and ADP-stimulated mitochondrial respiration in situ using saponin skinned muscle fibers. The in vitro maximal rates of respiration in isolated cardiac mitochondria from desmin-null and wild-type mice were similar. However, mitochondrial respiration in situ is significantly altered in desmin-null muscle. Both the maximal rate of ADP-stimulated oxygen consumption and the dissociation constant (K(m)) for ADP are significantly reduced in desmin-null cardiac and soleus muscle compared with controls. Respiratory parameters for desmin-null fast twitch gastrocnemius muscle were unaffected. Additionally, respiratory measurements in the presence of creatine indicate that coupling of creatine kinase and the adenine translocator is lost in desmin-null soleus muscle. This coupling is unaffected in cardiac muscle from desmin-null animals. All of these studies indicate that desmin IFs play a significant role in mitochondrial positioning and respiratory function in cardiac and skeletal muscle.
Figures








References
-
- Almahbobi G., Williams L.J., Han X.G., Hall P.F. Binding of lipid droplets and mitochondria to intermediate filaments in rat Leydig cells. J. Reprod. Fertil. 1993;98:209–217. - PubMed
-
- Bereiter-Hahn J., Voth M. Dynamics of mitochondria in living cellsshape changes, dislocations, fusion, and fission of mitochondria. Microsc. Res. Tech. 1994;27:198–219. - PubMed
-
- Bilak S.R., Sernett S.W., Bilak M.M., Bellin R.M., Stromer M.H., Huiatt T.W., Robson R.M. Properties of the novel intermediate filament protein synemin and its identification in mammalian muscle. Arch. Biochem. Biophys. 1998;355:63–76. - PubMed
-
- Capetanaki Y., Milner D. Desmin cytoskeleton in muscle integrity and function. Subcell. Biochem. 1998;31:463–495. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
