

New features of connectivity in piriform cortex visualized by intracellular injection of pyramidal cells suggest that "primary" olfactory cortex functions like "association" cortex in other sensory systems
- PMID: 10995842
- PMCID: PMC6772836
- DOI: 10.1523/JNEUROSCI.20-18-06974.2000
New features of connectivity in piriform cortex visualized by intracellular injection of pyramidal cells suggest that "primary" olfactory cortex functions like "association" cortex in other sensory systems
Abstract
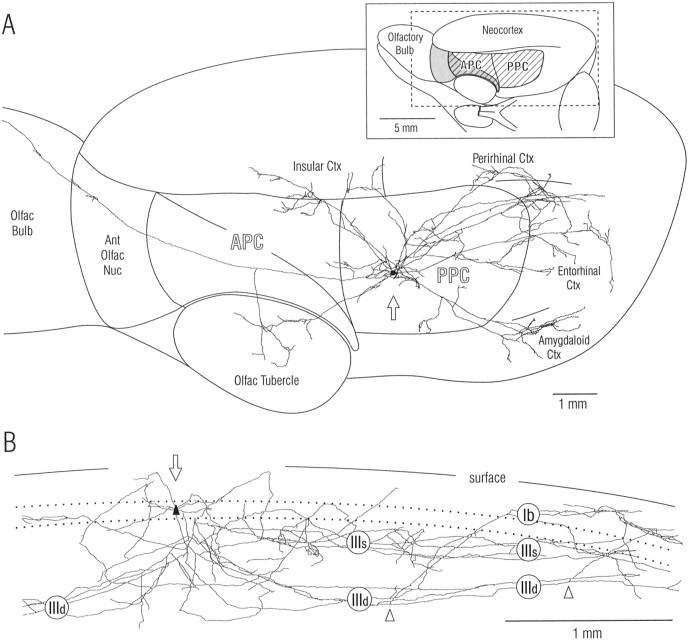
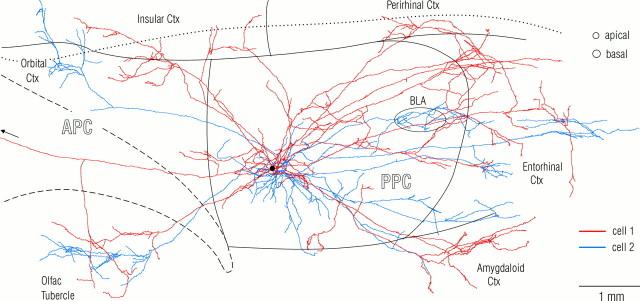
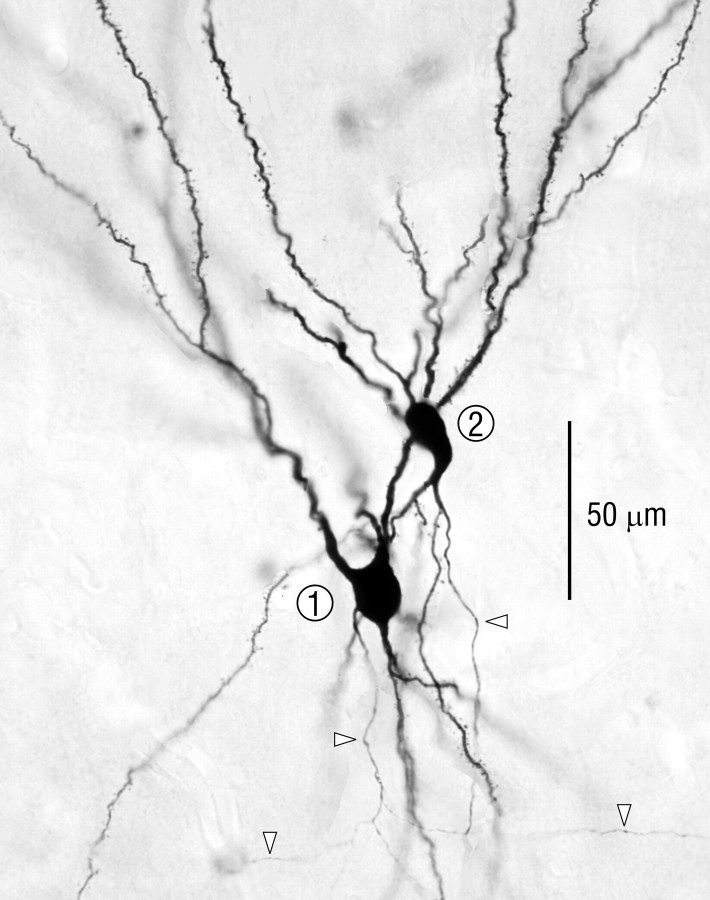
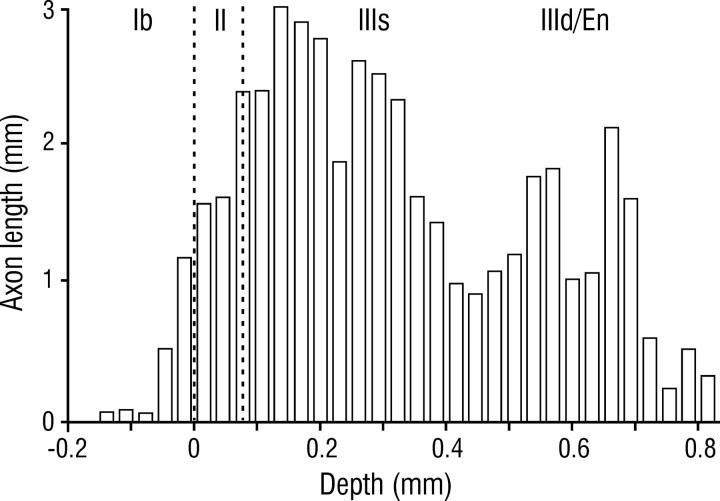
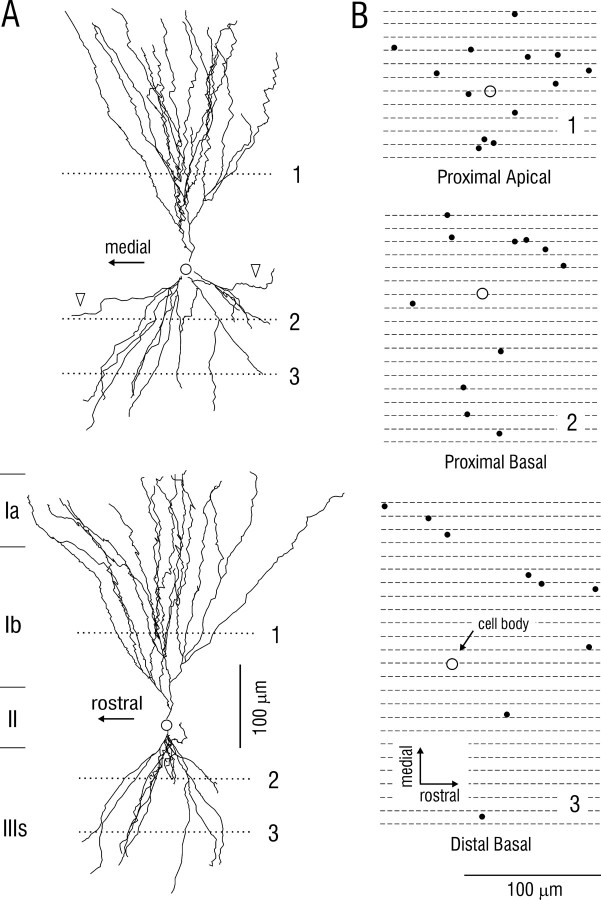
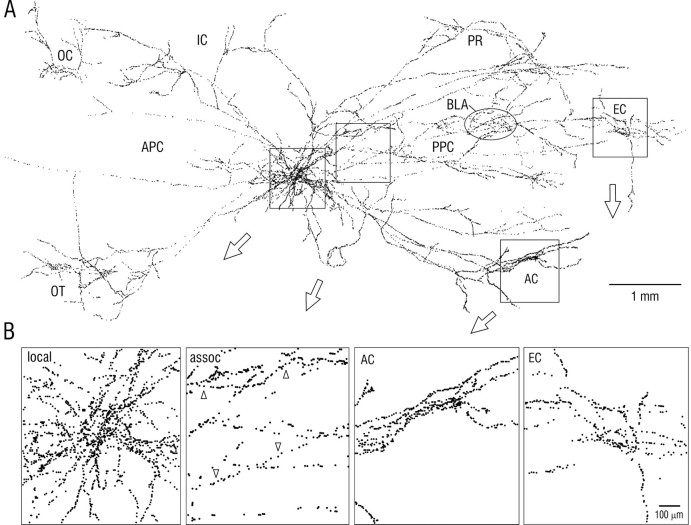
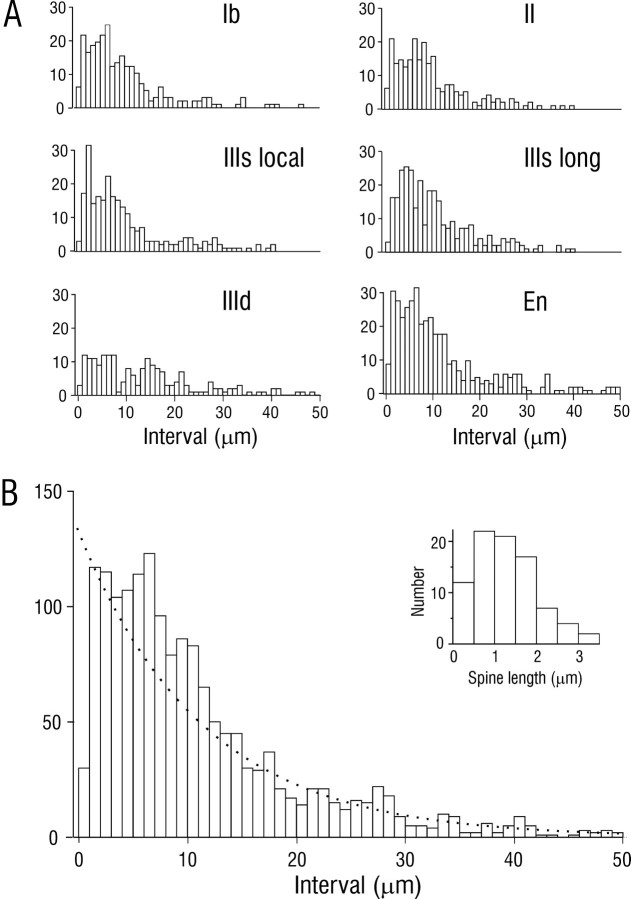
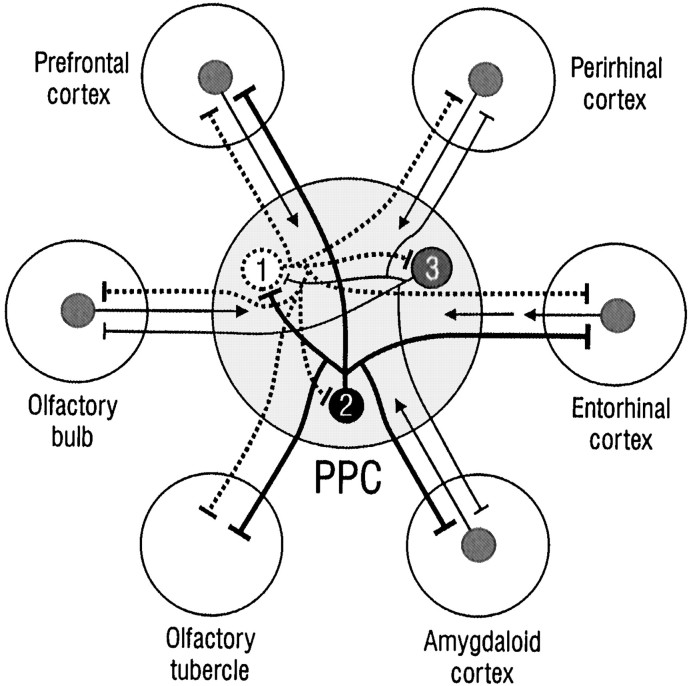
Associational connections of pyramidal cells in rat posterior piriform cortex were studied by direct visualization of axons stained by intracellular injection in vivo. The results revealed that individual cells have widespread axonal arbors that extend over nearly the full length of the cerebral hemisphere. Within piriform cortex these arbors are highly distributed with no regularly arranged patchy concentrations like those associated with the columnar organization in other primary sensory areas (i.e., where periodically arranged sets of cells have common response properties, inputs, and outputs). A lack of columnar organization was also indicated by a marked disparity in the intrinsic projection patterns of neighboring injected cells. Analysis of axonal branching patterns, bouton distributions, and dendritic arbors suggested that each pyramidal cell makes a small number of synaptic contacts on a large number (>1000) of other cells in piriform cortex at disparate locations. Axons from individual pyramidal cells also arborize extensively within many neighboring cortical areas, most of which send strong projections back to piriform cortex. These include areas involved in high-order functions in prefrontal, amygdaloid, entorhinal, and perirhinal cortex, to which there are few projections from other primary sensory areas. Our results suggest that piriform cortex performs correlative functions analogous to those in association areas of neocortex rather than those typical of primary sensory areas with which it has been traditionally classed. Findings from other studies suggest that the olfactory bulb subserves functions performed by primary areas in other sensory systems.
Figures
References
-
- Buck LB. Information coding in the vertebrate olfactory system. Annu Rev Neurosci. 1999;19:517–544. - PubMed
-
- Burkhalter A, Charles V. Organization of local axon collaterals of efferent projection neurons in rat visual cortex. J Comp Neurol. 1990;302:920–934. - PubMed
-
- Burwell RD, Amaral DG. Cortical afferents of the perirhinal, postrhinal, and entorhinal cortices of the rat. J Comp Neurol. 1998;398:179–205. - PubMed
-
- Chapin JK, Sadeq M, Guise JL. Corticocortical connections within the primary somatosensory cortex of the rat. J Comp Neurol. 1987;263:326–346. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources