

The role of identified neurotransmitter systems in the response of insular cortex to unfamiliar taste: activation of ERK1-2 and formation of a memory trace
- PMID: 10995847
- PMCID: PMC6772803
- DOI: 10.1523/JNEUROSCI.20-18-07017.2000
The role of identified neurotransmitter systems in the response of insular cortex to unfamiliar taste: activation of ERK1-2 and formation of a memory trace
Abstract
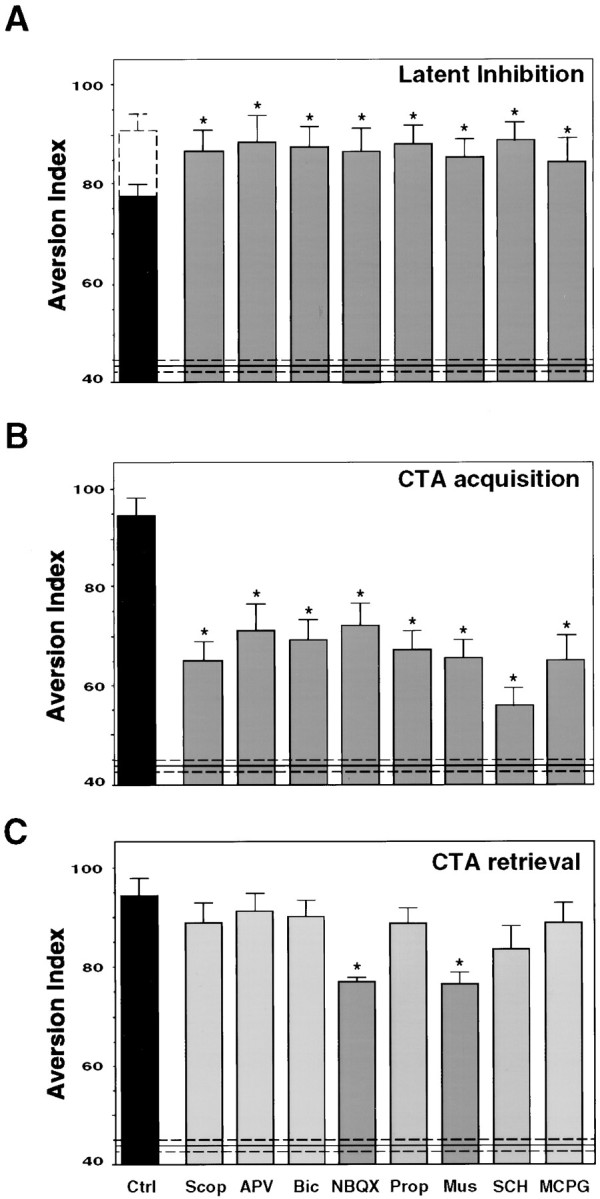
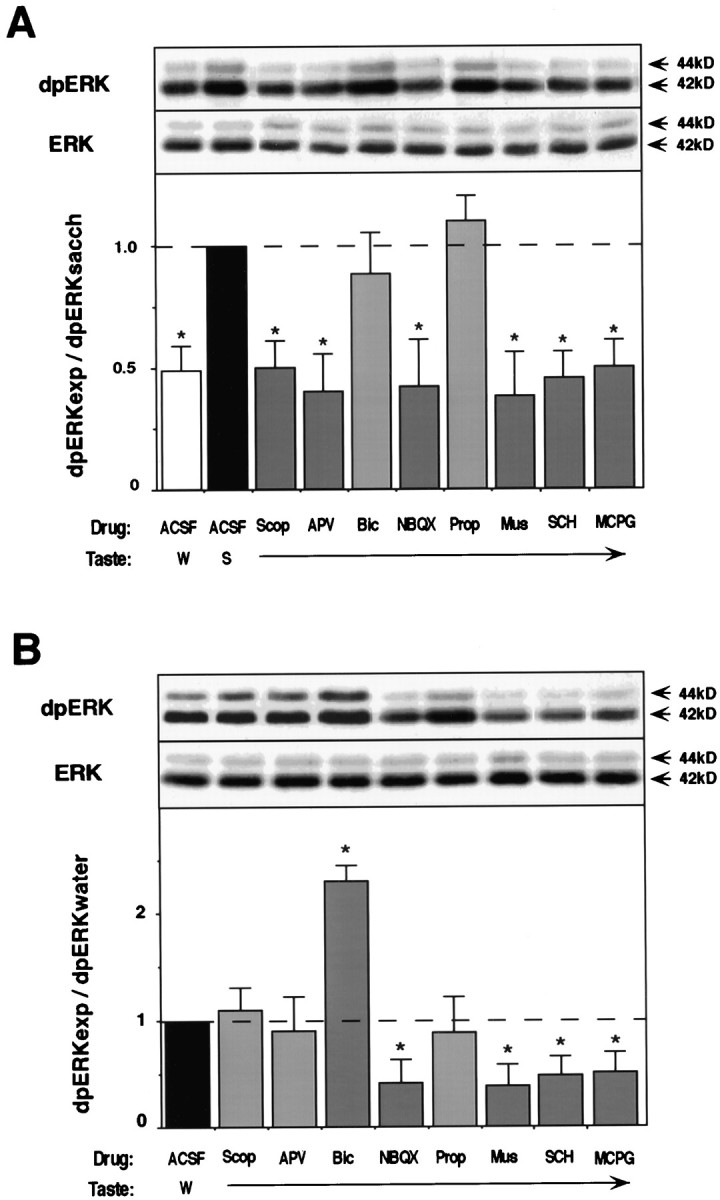
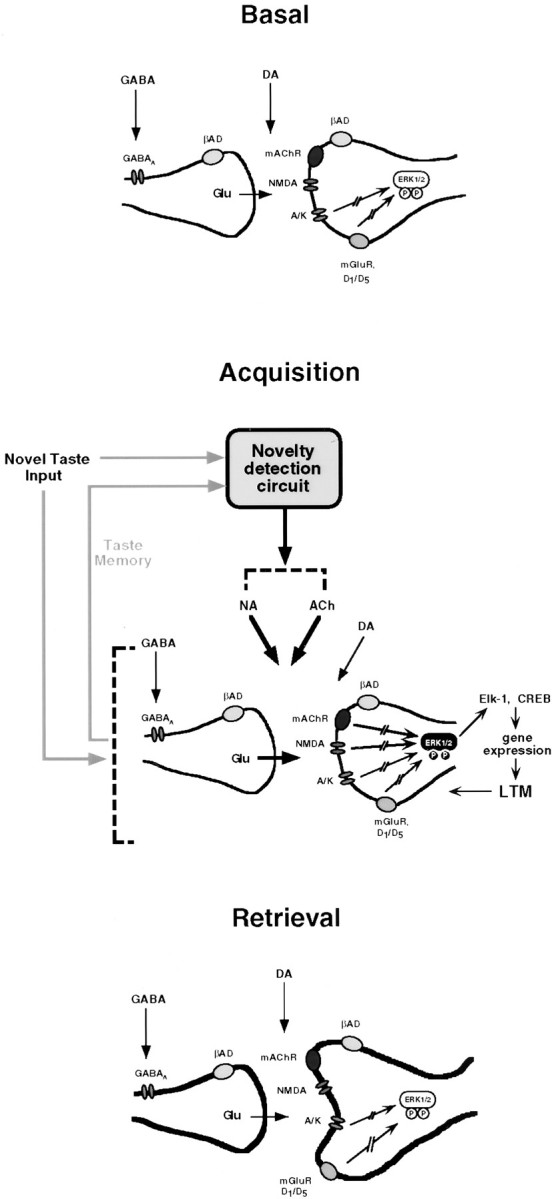
In the behaving rat, the consumption of an unfamiliar taste activates the extracellular signal-regulated kinase 1-2 (ERK1-2) in the insular cortex, which contains the taste cortex. In contrast, consumption of a familiar taste has no effect. Furthermore, activation of ERK1-2, culminating in modulation of gene expression, is obligatory for the encoding of long-term, but not short-term, memory of the new taste (Berman et al., 1998). Which neurotransmitter and neuromodulatory systems are involved in the activation of ERK1-2 by the unfamiliar taste and in the long-term encoding of the new taste information? Here we show, by the use of local microinjections of pharmacological agents to the insular cortex in the behaving rat, that multiple neurotransmitters and neuromodulators are required for encoding of taste memory in cortex. However, these systems vary in the specificity of their role in memory acquisition and in their contribution to the activation of ERK1-2. NMDA receptors, metabotropic glutamate receptors, muscarinic, and beta-adrenergic and dopaminergic receptors, all contribute to the acquisition of the new taste memory but not to its retrieval. Among these, only NMDA and muscarinic receptors specifically mediate taste-dependent activation of ERK1-2, whereas the beta-adrenergic function is independent of ERK1-2, and dopaminergic receptors regulate also the basal level of ERK1-2 activation. The data are discussed in the context of postulated novelty detection circuits in the central taste system.
Figures
References
-
- Ahissar E. Temporal-code to rate-code conversion by neuronal phase-locked loops. Neural Comput. 1998;10:597–650. - PubMed
-
- Asin KE, Montana WE. Studies on D1 and D2 dopamine receptor involvement in conditioned taste aversions. Pharmacol Biochem Behav. 1989;32:1033–1041. - PubMed
-
- Atkins CM, Selcher JC, Petraitis JJ, Trzaskos JM, Sweatt JD. The MAPK cascade is required for mammalian associative learning. Nat Neurosci. 1998;1:602–609. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous