

Long terminal repeat regions from exogenous but not endogenous feline leukemia viruses transactivate cellular gene expression
- PMID: 11000248
- PMCID: PMC112408
- DOI: 10.1128/jvi.74.20.9742-9748.2000
Long terminal repeat regions from exogenous but not endogenous feline leukemia viruses transactivate cellular gene expression
Abstract
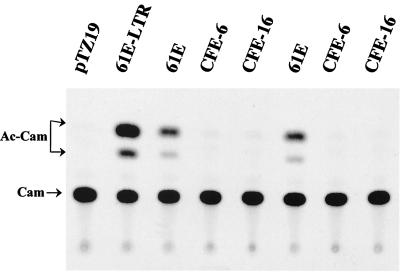
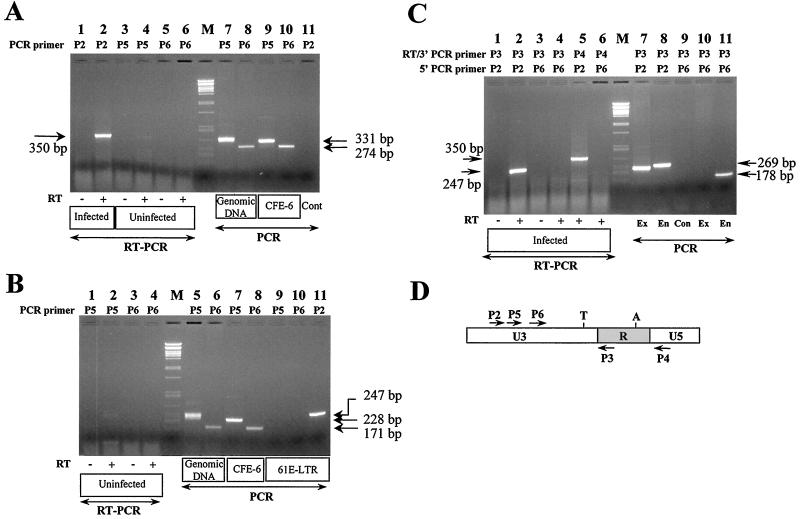
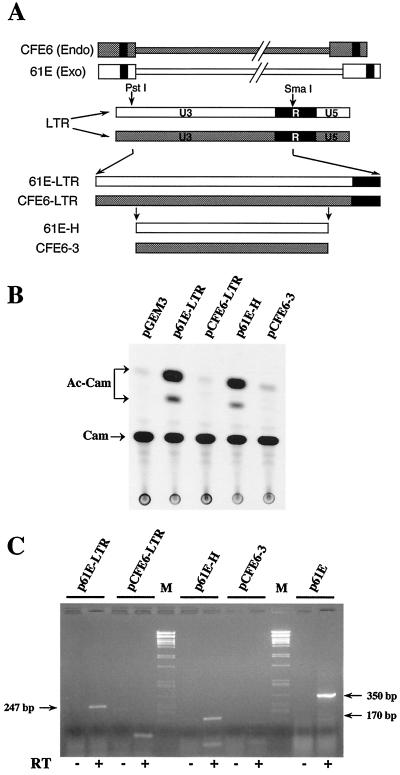
We have previously reported that the long terminal repeat (LTR) region of feline leukemia viruses (FeLVs) can enhance expression of certain cellular genes such as the collagenase IV gene and MCP-1 in trans (S. K. Ghosh and D. V. Faller, J. Virol. 73:4931-4940, 1999). Genomic DNA of all healthy feline species also contains LTR-like sequences that are related to exogenous FeLV LTRs. In this study, we evaluated the cellular gene transactivational potential of these endogenous FeLV LTR sequences. Unlike their exogenous FeLV counterparts, neither nearly full-length endogenous FeLV molecular clones (CFE-6 and CFE-16) nor their isolated LTRs were able to activate collagenase IV gene or MCP-1 expression in transient transfection assays. We had also demonstrated previously that production of an RNA transcript from exogenous FeLV LTRs correlates with their transactivational activity. In the present study, we demonstrate that the endogenous FeLV LTRs do not generate LTR-specific RNA transcripts in the feline embryo fibroblast cell line AH927. Furthermore, infection of AH927 cells by an exogenous FeLV subgroup A virus did not induce production of such LTR-specific transcripts from the endogenous proviral genomes, although the LTR-specific transcripts from the exogenous virus were readily detected. Finally, LTR-specific transcripts were not generated in BALB/3T3 cells transiently transfected with isolated CFE-6 LTR, in contrast to transfections with LTRs from exogenous viruses. Our data thus suggest that the inability of endogenous FeLV LTRs in gene transactivation is not due to cell line specificity or presence of any upstream inhibitory cis-acting element. Endogenous, nonleukemogenic FeLV LTRs, therefore, do not transactivate cellular gene expression, and this property appears to be specific to exogenous, leukemogenic FeLVs.
Figures

Similar articles
-
Feline leukemia virus long terminal repeat activates collagenase IV gene expression through AP-1.J Virol. 1999 Jun;73(6):4931-40. doi: 10.1128/JVI.73.6.4931-4940.1999. J Virol. 1999. PMID: 10233955 Free PMC article.
-
Mutations that abrogate transactivational activity of the feline leukemia virus long terminal repeat do not affect virus replication.Virology. 2003 May 10;309(2):294-305. doi: 10.1016/s0042-6822(03)00069-2. Virology. 2003. PMID: 12758176
-
Endogenous Feline Leukemia Virus (FeLV) siRNA Transcription May Interfere with Exogenous FeLV Infection.J Virol. 2021 Nov 9;95(23):e0007021. doi: 10.1128/JVI.00070-21. Epub 2021 Sep 8. J Virol. 2021. PMID: 34495702 Free PMC article.
-
Endogenous env elements: partners in generation of pathogenic feline leukemia viruses.Virus Genes. 1995;11(2-3):147-61. doi: 10.1007/BF01728655. Virus Genes. 1995. PMID: 8828142 Review.
-
Quantification of endogenous and exogenous feline leukemia virus sequences by real-time PCR assays.Vet Immunol Immunopathol. 2008 May 15;123(1-2):129-33. doi: 10.1016/j.vetimm.2008.01.027. Epub 2008 Jan 19. Vet Immunol Immunopathol. 2008. PMID: 18295344 Review.
Cited by
-
High prevalence of non-productive FeLV infection in necropsied cats and significant association with pathological findings.Vet Immunol Immunopathol. 2010 Jul;136(1-2):71-80. doi: 10.1016/j.vetimm.2010.02.014. Epub 2010 Mar 25. Vet Immunol Immunopathol. 2010. PMID: 20398945 Free PMC article.
-
Leukemia virus long terminal repeat activates NFkappaB pathway by a TLR3-dependent mechanism.Virology. 2006 Feb 20;345(2):390-403. doi: 10.1016/j.virol.2005.10.003. Epub 2005 Nov 14. Virology. 2006. PMID: 16289658 Free PMC article.
-
Identification of LTR-specific small non-coding RNA in FeLV infected cells.FEBS Lett. 2009 Apr 17;583(8):1386-90. doi: 10.1016/j.febslet.2009.03.056. Epub 2009 Mar 29. FEBS Lett. 2009. PMID: 19336234 Free PMC article.
-
Prevalence of feline leukaemia provirus DNA in feline lymphomas.J Feline Med Surg. 2010 Dec;12(12):929-35. doi: 10.1016/j.jfms.2010.07.006. Epub 2010 Oct 30. J Feline Med Surg. 2010. PMID: 21036089 Free PMC article.
-
Endogenous feline leukemia virus long terminal repeat integration site diversity is highly variable in related and unrelated domestic cats.Retrovirology. 2024 Feb 12;21(1):3. doi: 10.1186/s12977-024-00635-0. Retrovirology. 2024. PMID: 38347535 Free PMC article.
References
-
- Benveniste R E, Sherr C J, Todaro G J. Evolution of type C viral genes: origin of feline leukemia virus. Science. 1975;190:886–888. - PubMed
-
- Boeke J D, Stoye J P. Retrotransposons, endogenous retroviruses, and the evolution of retroelements. In: Coffin J M, Hughes S H, Varmus H E, editors. Retroviruses. Cold Spring Harbor, N.Y: Cold Spring Harbor Laboratory Press; 1997. pp. 343–436. - PubMed
-
- Busch M P, Devi B G, Soe L H, Perbal B, Baluda M A, Roy-Burman P. Characterization of the expression of cellular retrovirus genes and oncogenes in feline cells. Hematol Oncol. 1983;1:61–75. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
