

Alphavirus RNA genome repair and evolution: molecular characterization of infectious sindbis virus isolates lacking a known conserved motif at the 3' end of the genome
- PMID: 11000254
- PMCID: PMC112414
- DOI: 10.1128/jvi.74.20.9776-9785.2000
Alphavirus RNA genome repair and evolution: molecular characterization of infectious sindbis virus isolates lacking a known conserved motif at the 3' end of the genome
Abstract
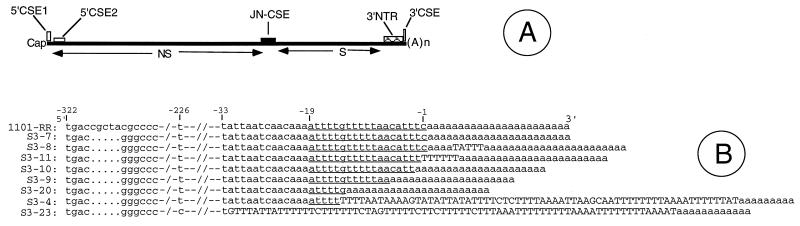
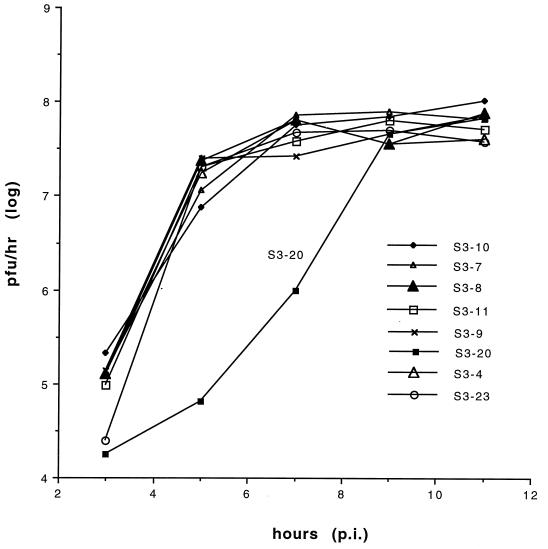
The 3' nontranslated region of the genomes of Sindbis virus (SIN) and other alphaviruses carries several repeat sequence elements (RSEs) as well as a 19-nucleotide (nt) conserved sequence element (3'CSE). The 3'CSE and the adjoining poly(A) tail of the SIN genome are thought to act as viral promoters for negative-sense RNA synthesis and genome replication. Eight different SIN isolates that carry altered 3'CSEs were studied in detail to evaluate the role of the 3'CSE in genome replication. The salient findings of this study as it applies to SIN infection of BHK cells are as follows: i) the classical 19-nt 3'CSE of the SIN genome is not essential for genome replication, long-term stability, or packaging; ii) compensatory amino acid or nucleotide changes within the SIN genomes are not required to counteract base changes in the 3' terminal motifs of the SIN genome; iii) the 5' 1-kb regions of all SIN genomes, regardless of the differences in 3' terminal motifs, do not undergo any base changes even after 18 passages; iv) although extensive addition of AU-rich motifs occurs in the SIN genomes carrying defective 3'CSE, these are not essential for genome viability or function; and v) the newly added AU-rich motifs are composed predominantly of RSEs. These findings are consistent with the idea that the 3' terminal AU-rich motifs of the SIN genomes do not bind directly to the viral polymerase and that cellular proteins with broad AU-rich binding specificity may mediate this interaction. In addition to the classical 3'CSE, other RNA motifs located elsewhere in the SIN genome must play a major role in template selection by the SIN RNA polymerase.
Figures

Similar articles
-
In vivo addition of poly(A) tail and AU-rich sequences to the 3' terminus of the Sindbis virus RNA genome: a novel 3'-end repair pathway.J Virol. 1999 Mar;73(3):2410-9. doi: 10.1128/JVI.73.3.2410-2419.1999. J Virol. 1999. PMID: 9971825 Free PMC article.
-
Efficient replication, and evolution of Sindbis virus genomes with non-canonical 3'A/U-rich elements (NC3ARE) in neonatal mice.Virus Genes. 2007 Dec;35(3):651-62. doi: 10.1007/s11262-007-0130-z. Epub 2007 Jul 7. Virus Genes. 2007. PMID: 17616797
-
RNA-RNA recombination in Sindbis virus: roles of the 3' conserved motif, poly(A) tail, and nonviral sequences of template RNAs in polymerase recognition and template switching.J Virol. 1997 Apr;71(4):2693-704. doi: 10.1128/JVI.71.4.2693-2704.1997. J Virol. 1997. PMID: 9060622 Free PMC article.
-
Template requirements for recognition and copying by Sindbis virus RNA-dependent RNA polymerase.Virology. 2007 Feb 5;358(1):221-32. doi: 10.1016/j.virol.2006.08.022. Epub 2006 Sep 18. Virology. 2007. PMID: 16979682
-
Functions of the 3' and 5' genome RNA regions of members of the genus Flavivirus.Virus Res. 2015 Aug 3;206:108-19. doi: 10.1016/j.virusres.2015.02.006. Epub 2015 Feb 13. Virus Res. 2015. PMID: 25683510 Free PMC article. Review.
Cited by
-
Requirements at the 3' end of the sindbis virus genome for efficient synthesis of minus-strand RNA.J Virol. 2005 Apr;79(8):4630-9. doi: 10.1128/JVI.79.8.4630-4639.2005. J Virol. 2005. PMID: 15795249 Free PMC article.
-
Recent Advancement in mRNA Vaccine Development and Applications.Pharmaceutics. 2023 Jul 18;15(7):1972. doi: 10.3390/pharmaceutics15071972. Pharmaceutics. 2023. PMID: 37514158 Free PMC article. Review.
-
Selection of functional 5' cis-acting elements promoting efficient sindbis virus genome replication.J Virol. 2004 Jan;78(1):61-75. doi: 10.1128/jvi.78.1.61-75.2004. J Virol. 2004. PMID: 14671088 Free PMC article.
-
Identification of a novel Getah virus by Virus-Discovery-cDNA random amplified polymorphic DNA (RAPD).BMC Microbiol. 2012 Dec 27;12:305. doi: 10.1186/1471-2180-12-305. BMC Microbiol. 2012. PMID: 23268691 Free PMC article.
-
The 5' and 3' ends of alphavirus RNAs--Non-coding is not non-functional.Virus Res. 2015 Aug 3;206:99-107. doi: 10.1016/j.virusres.2015.01.016. Epub 2015 Jan 25. Virus Res. 2015. PMID: 25630058 Free PMC article. Review.
References
-
- Anant S, Davidson N O. An AU-rich sequence element (UUUN[A/U]U) downstream of the edited C in apolipoprotein B mRNA is a high-affinity binding site for Apobec-1: binding of Apobec-1 to this motif in the 3′ untranslated region of c-myc increases mRNA stability. Mol Cell Biol. 2000;20:1982–1992. - PMC - PubMed
-
- Benne R. RNA editing: an overview. In: Benne R, editor. RNA editing. Chichester, England: Ellis Horwood; 1993. pp. 13–24.
-
- Blackburn E H. Telomerase. In: Gesteland R F, Atkins J F, editors. The RNA world. Cold Spring Harbor, N.Y: Cold Spring Harbor Laboratory Press; 1993. pp. 557–576.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
