

Induction of human fetal globin gene expression by a novel erythroid factor, NF-E4
- PMID: 11003662
- PMCID: PMC86334
- DOI: 10.1128/MCB.20.20.7662-7672.2000
Induction of human fetal globin gene expression by a novel erythroid factor, NF-E4
Abstract
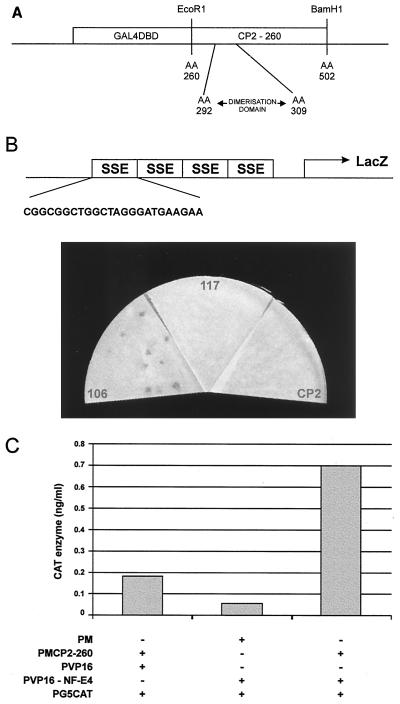
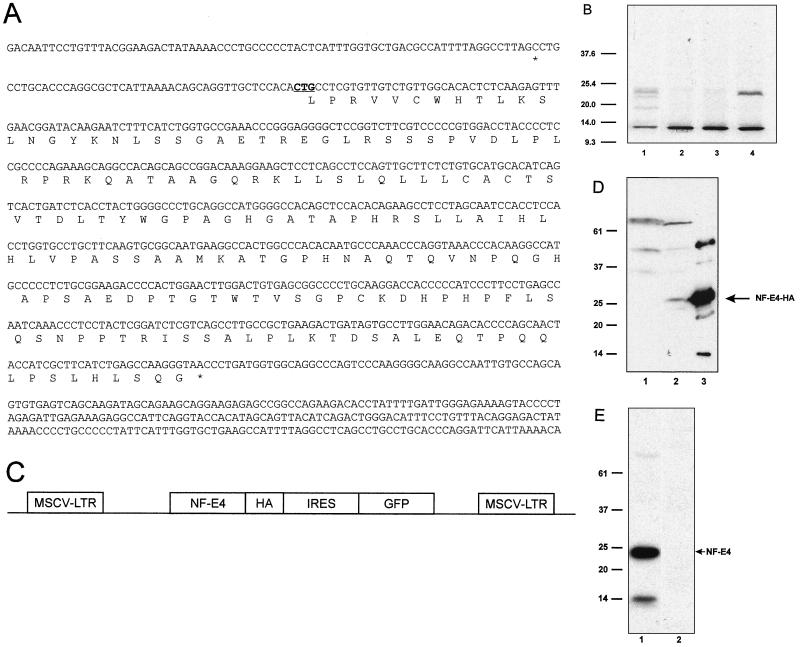
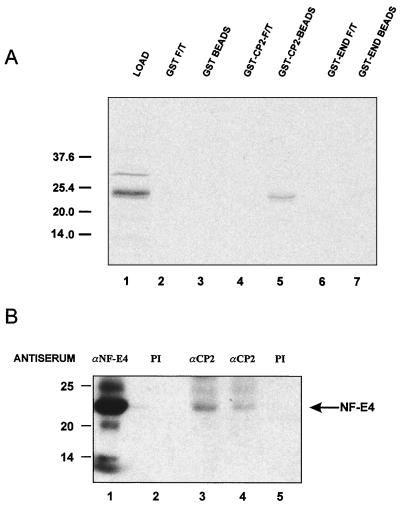
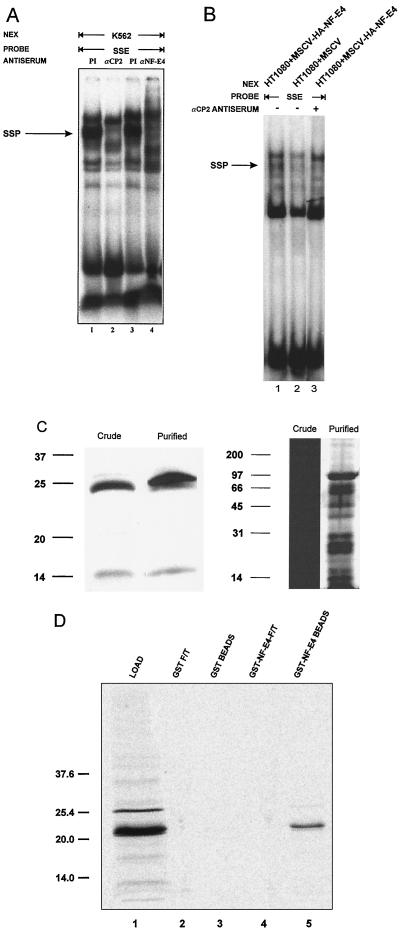
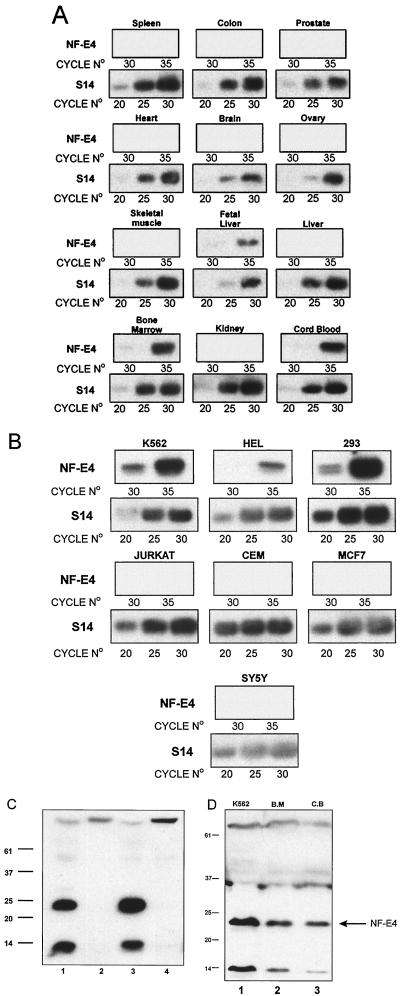
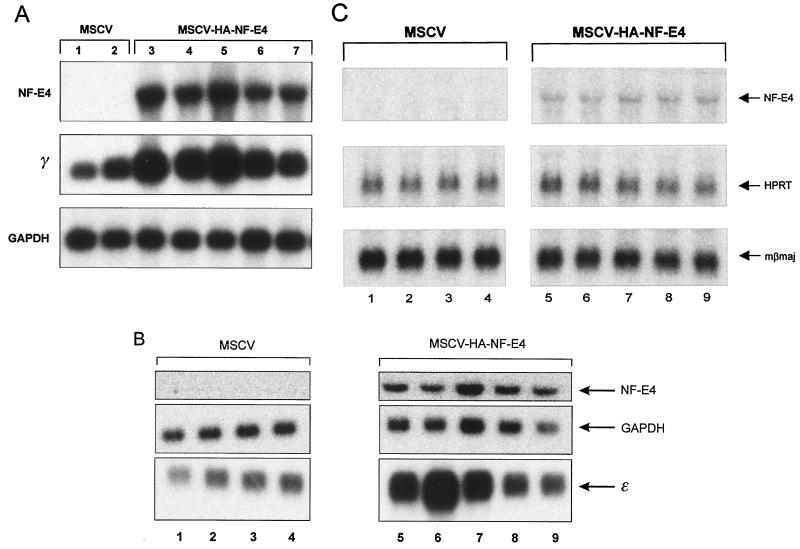
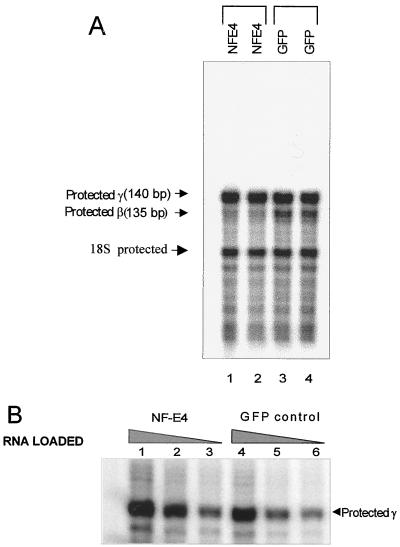
The stage selector protein (SSP) is a heteromeric complex involved in preferential expression of the human gamma-globin genes in fetal-erythroid cells. We have previously identified the ubiquitous transcription factor CP2 as a component of this complex. Using the protein dimerization domain of CP2 in a yeast two-hybrid screen, we have cloned a novel gene, NF-E4, encoding the tissue-restricted component of the SSP. NF-E4 and CP2 coimmunoprecipitate from extract derived from a fetal-erythroid cell line, and antiserum to NF-E4 ablates binding of the SSP to the gamma promoter. NF-E4 is expressed in fetal liver, cord blood, and bone marrow and in the K562 and HEL cell lines, which constitutively express the fetal globin genes. Enforced expression of NF-E4 in K562 cells and primary erythroid progenitors induces endogenous fetal globin gene expression, suggesting a possible strategy for therapeutic intervention in the hemoglobinopathies.
Figures
References
-
- Acland P, Dixon M, Peters G, Dickson C. The subcellular fate of the Int-2 oncoprotein is determined by choice of initiation codon. Nature. 1990;343:662–665. - PubMed
-
- Amrolia P J, Cunningham J M, Jane S M. Maximal activity of an erythroid-specific enhancer requires the presence of specific protein binding sites in linked promoters. J Biol Chem. 1998;273:13593–13598. - PubMed
-
- Armstrong J A, Bieker J J, Emerson B M. A SWI/SNF-related chromatin remodelling complex, E-RC1, is required for tissue-specific transcriptional regulation by EKLF in vitro. Cell. 1998;95:93–104. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases