

Inhibition of ErbB-2 mitogenic and transforming activity by RALT, a mitogen-induced signal transducer which binds to the ErbB-2 kinase domain
- PMID: 11003669
- PMCID: PMC86354
- DOI: 10.1128/MCB.20.20.7735-7750.2000
Inhibition of ErbB-2 mitogenic and transforming activity by RALT, a mitogen-induced signal transducer which binds to the ErbB-2 kinase domain
Abstract
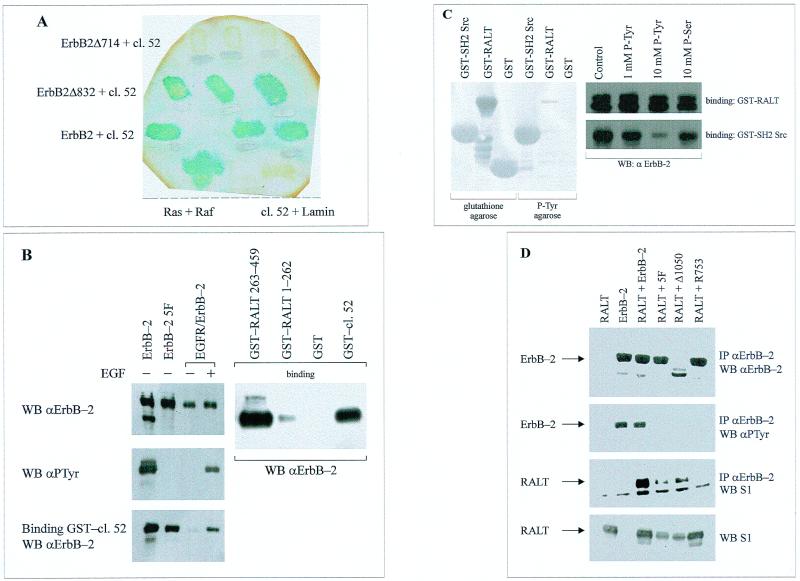
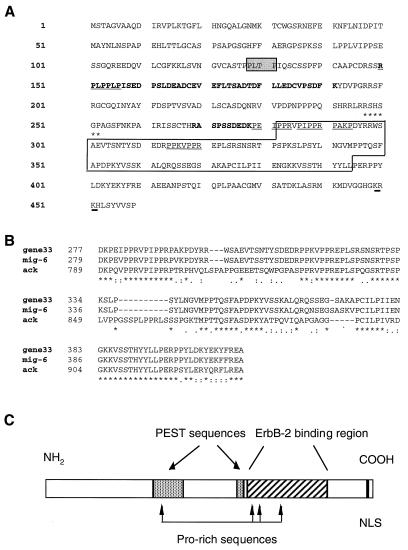
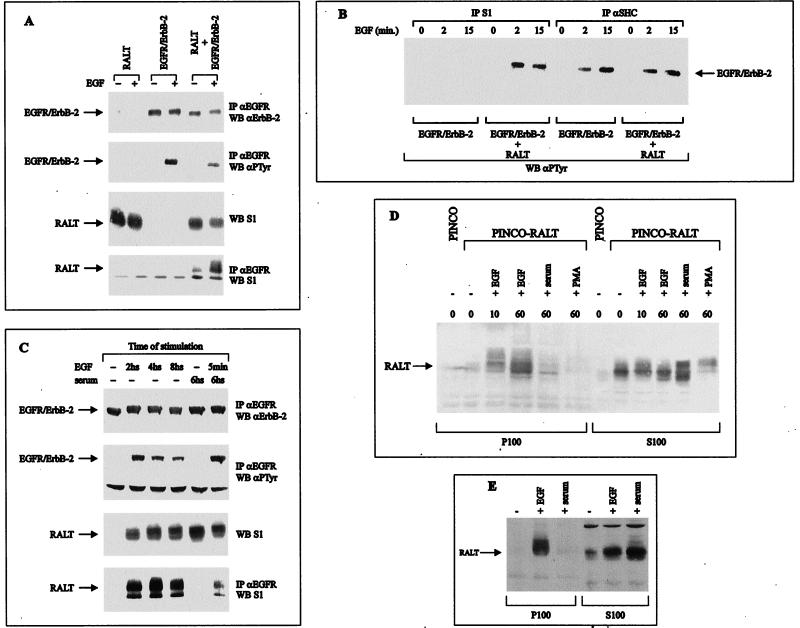
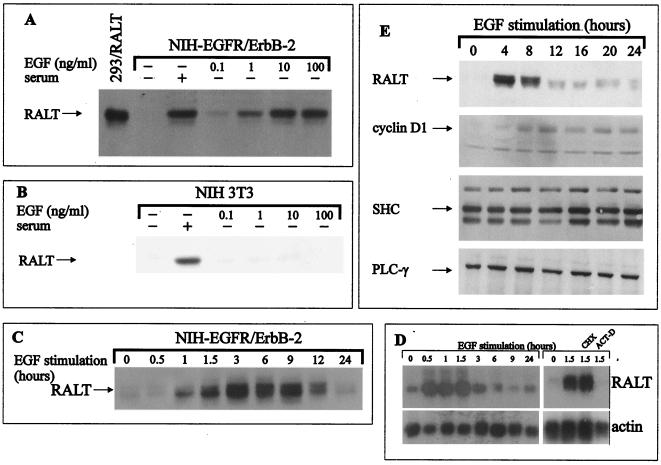
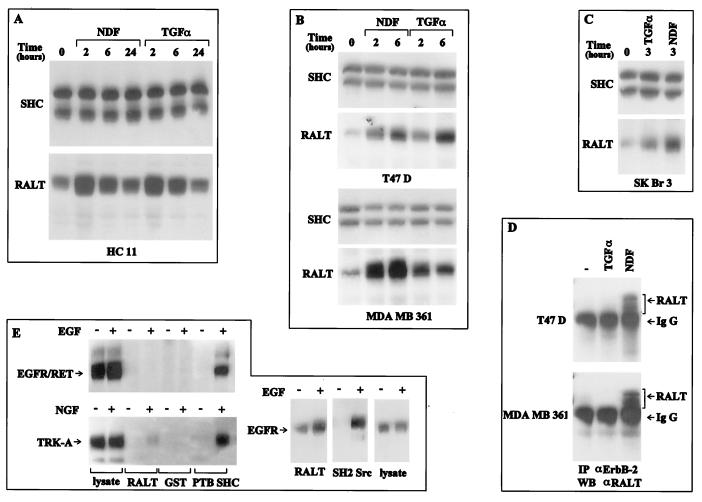
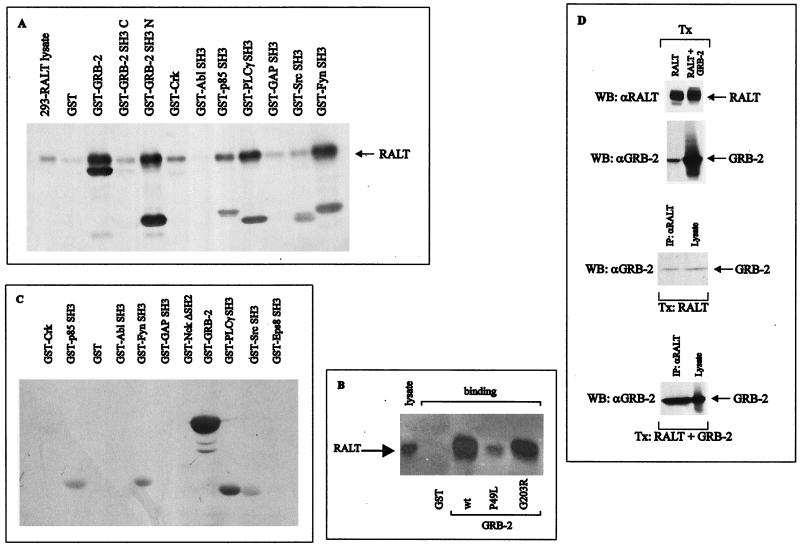
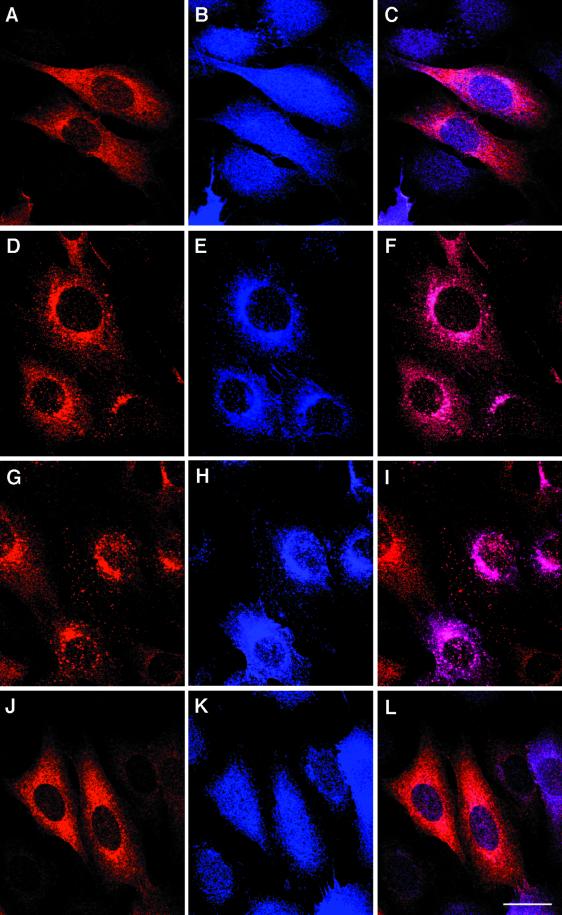
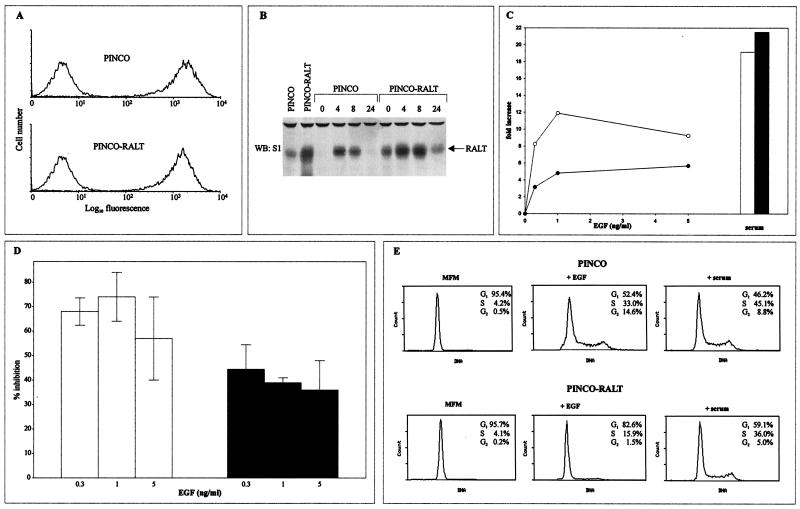
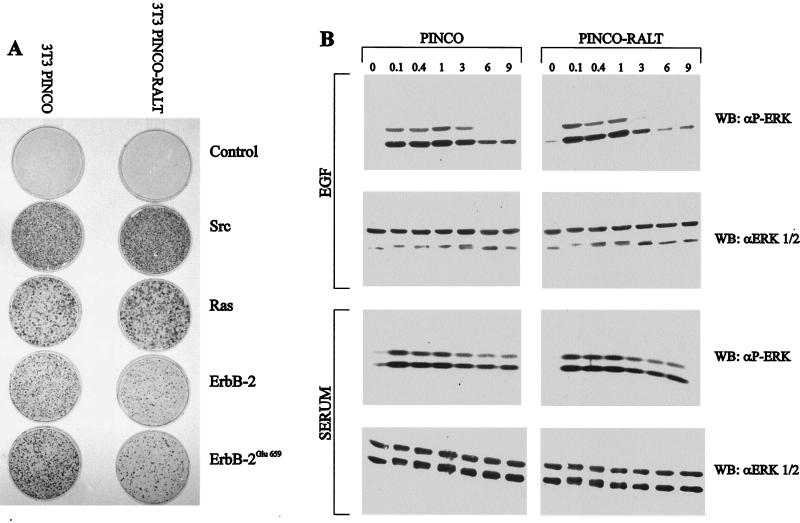
The product of rat gene 33 was identified as an ErbB-2-interacting protein in a two-hybrid screen employing the ErbB-2 juxtamembrane and kinase domains as bait. This interaction was reproduced in vitro with a glutathione S-transferase fusion protein spanning positions 282 to 395 of the 459-residue gene 33 protein. Activation of ErbB-2 catalytic function was required for ErbB-2-gene 33 physical interaction in living cells, whereas ErbB-2 autophosphorylation was dispensable. Expression of gene 33 protein was absent in growth-arrested NIH 3T3 fibroblasts but was induced within 60 to 90 min of serum stimulation or activation of the ErbB-2 kinase and decreased sharply upon entry into S phase. New differentiation factor stimulation of mitogen-deprived mammary epithelial cells also caused accumulation of gene 33 protein, which could be found in a complex with ErbB-2. Overexpression of gene 33 protein in mouse fibroblasts inhibited (i) cell proliferation driven by ErbB-2 but not by serum, (ii) cell transformation induced by ErbB-2 but not by Ras or Src, and (iii) sustained activation of ERK 1 and 2 by ErbB-2 but not by serum. The gene 33 protein may convey inhibitory signals downstream to ErbB-2 by virtue of its association with SH3-containing proteins, including GRB-2, which was found to associate with gene 33 protein in living cells. These data indicate that the gene 33 protein is a feedback inhibitor of ErbB-2 mitogenic function and a suppressor of ErbB-2 oncogenic activity. We propose that the gene 33 protein be renamed with the acronym RALT (receptor-associated late transducer).
Figures
References
-
- Alroy I, Yarden Y. The ErbB signaling network in embryogenesis and oncogenesis: signal diversification through combinatorial ligand-receptor interactions. FEBS Lett. 1997;410:83–86. - PubMed
-
- Anton E S, Marchionni M A, Lee K F, Rakic P. Role of GGF/neuregulin signaling in interactions between migrating neurons and radial glia in the developing cerebral cortex. Development. 1997;124:3501–3510. - PubMed
-
- Baulida J, Kraus M H, Alimandi M, Di Fiore P P, Carpenter G. All ErbB receptors other than the epidermal growth factor receptor are endocytosis impaired. J Biol Chem. 1996;271:5251–5257. - PubMed
-
- Beerli R R, Hynes N E. Epidermal growth factor-related peptides activate distinct subsets of ErbB receptors and differ in their biological activities. J Biol Chem. 1996;271:6071–6076. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous