

On the architecture of the gram-negative bacterial murein sacculus
- PMID: 11004199
- PMCID: PMC94722
- DOI: 10.1128/JB.182.20.5925-5930.2000
On the architecture of the gram-negative bacterial murein sacculus
Abstract
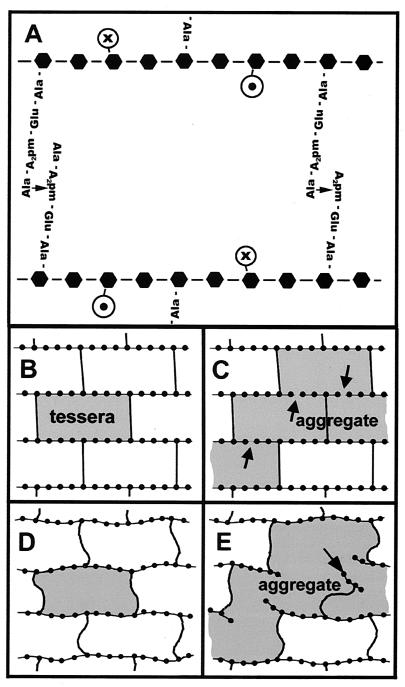
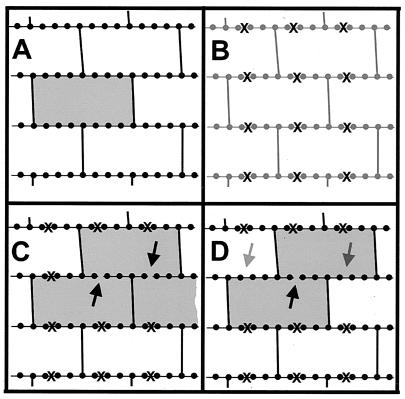
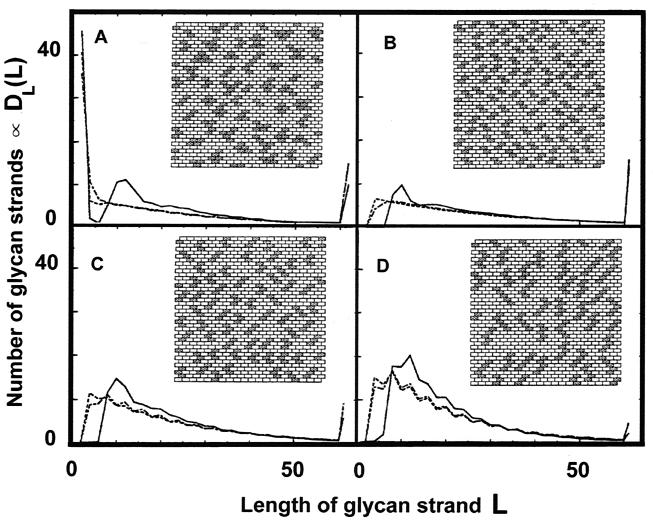
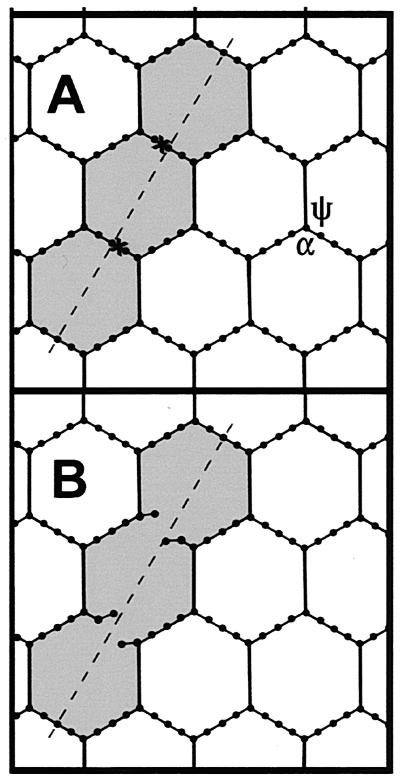
The peptidoglycan network of the murein sacculus must be porous so that nutrients, waste products, and secreted proteins can pass through. Using Escherichia coli and Pseudomonas aeruginosa as a baseline for gram-negative sacculi, the hole size distribution in the peptidoglycan network has been modeled by computer simulation to deduce the network's properties. By requiring that the distribution of glycan chain lengths predicted by the model be in accord with the distribution observed, we conclude that the holes are slits running essentially perpendicular to the local axis of the glycan chains (i. e., the slits run along the long axis of the cell). This result is in accord with previous permeability measurements of Beveridge and Jack and Demchik and Koch. We outline possible advantages that might accrue to the bacterium via this architecture and suggest ways in which such defect structures might be detected. Certainly, large molecules do penetrate the peptidoglycan layer of gram-negative bacteria, and the small slits that we suggest might be made larger by the bacterium.
Figures
Similar articles
-
The architecture of the murein (peptidoglycan) in gram-negative bacteria: vertical scaffold or horizontal layer(s)?J Bacteriol. 2004 Sep;186(18):5978-87. doi: 10.1128/JB.186.18.5978-5987.2004. J Bacteriol. 2004. PMID: 15342566 Free PMC article. Review. No abstract available.
-
Thickness and elasticity of gram-negative murein sacculi measured by atomic force microscopy.J Bacteriol. 1999 Nov;181(22):6865-75. doi: 10.1128/JB.181.22.6865-6875.1999. J Bacteriol. 1999. PMID: 10559150 Free PMC article.
-
Tertiary structure of bacterial murein: the scaffold model.J Bacteriol. 2003 Jun;185(11):3458-68. doi: 10.1128/JB.185.11.3458-3468.2003. J Bacteriol. 2003. PMID: 12754246 Free PMC article.
-
Murein (peptidoglycan) structure, architecture and biosynthesis in Escherichia coli.Biochim Biophys Acta. 2008 Sep;1778(9):1714-34. doi: 10.1016/j.bbamem.2007.06.007. Epub 2007 Jun 16. Biochim Biophys Acta. 2008. PMID: 17658458 Review.
-
Molecular organization of Gram-negative peptidoglycan.Proc Natl Acad Sci U S A. 2008 Dec 2;105(48):18953-7. doi: 10.1073/pnas.0808035105. Epub 2008 Nov 24. Proc Natl Acad Sci U S A. 2008. PMID: 19033194 Free PMC article.
Cited by
-
Escherichia coli peptidoglycan structure and mechanics as predicted by atomic-scale simulations.PLoS Comput Biol. 2014 Feb 20;10(2):e1003475. doi: 10.1371/journal.pcbi.1003475. eCollection 2014 Feb. PLoS Comput Biol. 2014. PMID: 24586129 Free PMC article.
-
The Escherichia coli Outer Membrane β-Barrel Assembly Machinery (BAM) Anchors the Peptidoglycan Layer by Spanning It with All Subunits.Int J Mol Sci. 2021 Feb 12;22(4):1853. doi: 10.3390/ijms22041853. Int J Mol Sci. 2021. PMID: 33673366 Free PMC article.
-
The architecture of the murein (peptidoglycan) in gram-negative bacteria: vertical scaffold or horizontal layer(s)?J Bacteriol. 2004 Sep;186(18):5978-87. doi: 10.1128/JB.186.18.5978-5987.2004. J Bacteriol. 2004. PMID: 15342566 Free PMC article. Review. No abstract available.
-
Cryo-transmission electron microscopy of frozen-hydrated sections of Escherichia coli and Pseudomonas aeruginosa.J Bacteriol. 2003 Oct;185(20):6112-8. doi: 10.1128/JB.185.20.6112-6118.2003. J Bacteriol. 2003. PMID: 14526023 Free PMC article.
-
Cell wall peptidoglycan in Mycobacterium tuberculosis: An Achilles' heel for the TB-causing pathogen.FEMS Microbiol Rev. 2019 Sep 1;43(5):548-575. doi: 10.1093/femsre/fuz016. FEMS Microbiol Rev. 2019. PMID: 31183501 Free PMC article. Review.
References
-
- Beveridge T J, Popham T J, Cole R M. Electron microscopy. In: Gerhardt P, editor. Methods for general and molecular bacteriology. Washington, D.C.: American Society for Microbiology; 1994. pp. 42–71.
-
- Binder K, editor. Applications of the Monte Carlo method in statistical physics. Heidelberg, Germany: Springer-Verlag; 1984.
-
- de Jonge B. Peptidoglycan synthesis during the cell cycle of Escherichia coli: structure and mode of insertion. Ph.D. thesis. Amsterdam, The Netherlands: University of Amsterdam; 1989. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
