

SOS3 function in plant salt tolerance requires N-myristoylation and calcium binding
- PMID: 11006339
- PMCID: PMC149077
- DOI: 10.1105/tpc.12.9.1667
SOS3 function in plant salt tolerance requires N-myristoylation and calcium binding
Abstract
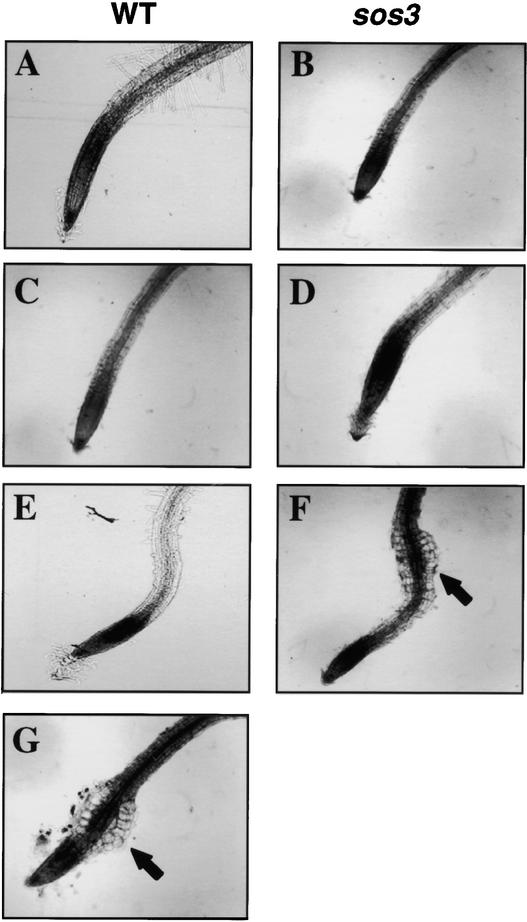
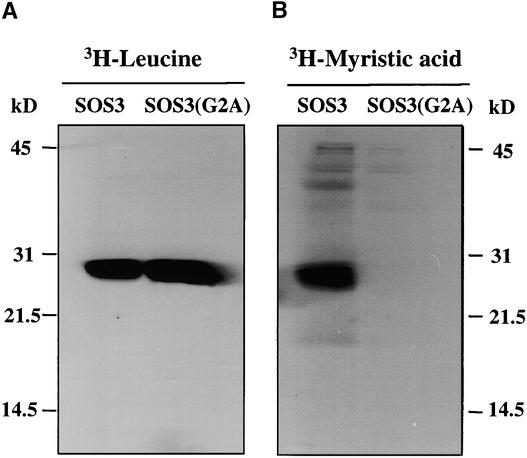
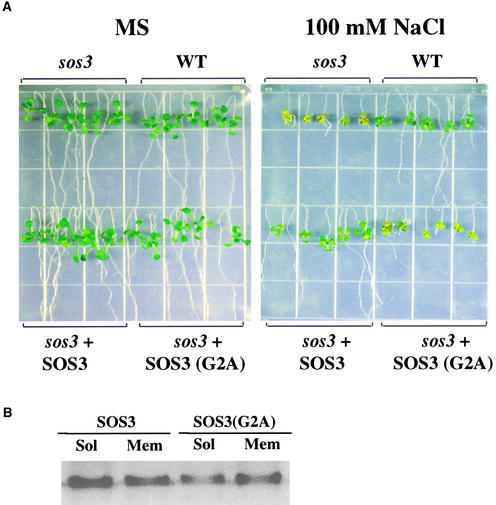
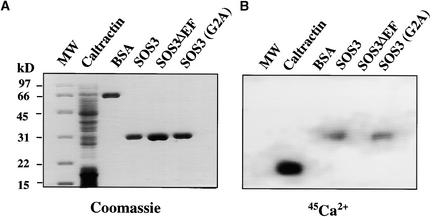
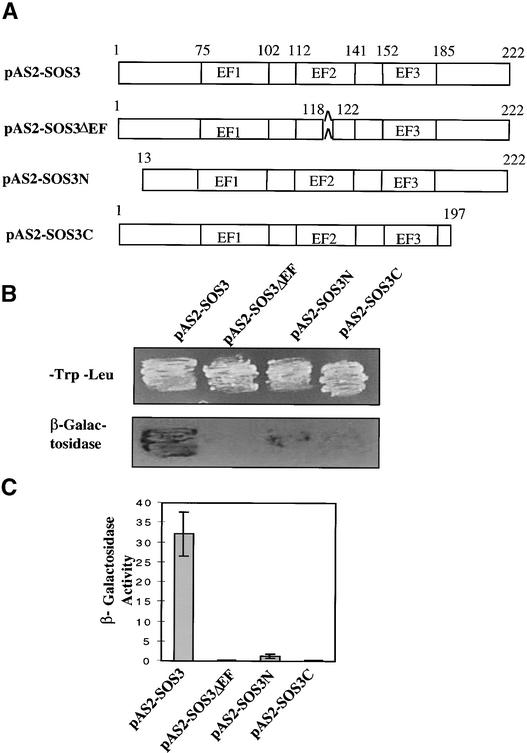
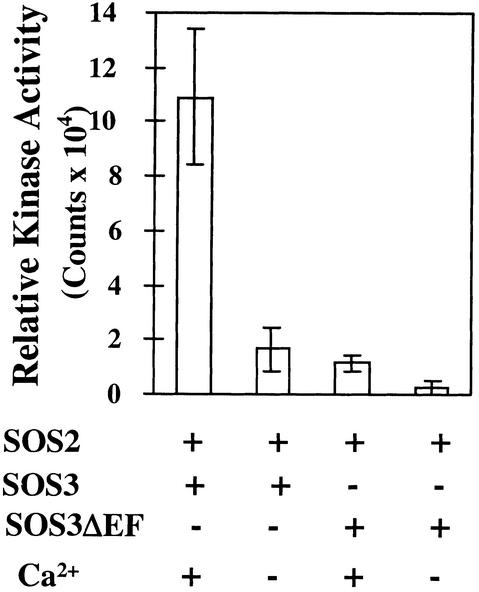
The salt tolerance gene SOS3 (for salt overly sensitive3) of Arabidopsis is predicted to encode a calcium binding protein with an N-myristoylation signature sequence. Here, we examine the myristoylation and calcium binding properties of SOS3 and their functional significance in plant tolerance to salt. Treatment of young Arabidopsis seedlings with the myristoylation inhibitor 2-hydroxymyristic acid caused the swelling of root tips, mimicking the phenotype of the salt-hypersensitive mutant sos3-1. In vitro translation assays with reticulocyte showed that the SOS3 protein was myristoylated. Targeted mutagenesis of the N-terminal glycine-2 to alanine prevented the myristoylation of SOS3. The functional significance of SOS3 myristoylation was examined by expressing the wild-type myristoylated SOS3 and the mutated nonmyristoylated SOS3 in the sos3-1 mutant. Expression of the myristoylated but not the nonmyristoylated SOS3 complemented the salt-hypersensitive phenotype of sos3-1 plants. No significant difference in membrane association was observed between the myristoylated and nonmyristoylated SOS3. Gel mobility shift and (45)Ca(2)+ overlay assays demonstrated that SOS3 is a unique calcium binding protein and that the sos3-1 mutation substantially reduced the capacity of SOS3 to bind calcium. The resulting mutant SOS3 protein was not able to interact with the SOS2 protein kinase and was less capable of activating it. Together, these results strongly suggest that both N-myristoylation and calcium binding are required for SOS3 function in plant salt tolerance.
Figures


References
-
- Ames, J.B., Ishima, R., Tanaka, T., Gordon, J.I., Stryer, L., and Ikura, M. (1997). Molecular mechanisms of calcium-myristoyl switches. Nature 389, 198–202. - PubMed
-
- Dizhoor, A.M., Chen, C.K., Olshevskaya, E., Sinelnikova, V.V., Philipov, P., and Hurley, J.B. (1993). Role of acylated amino terminus of recoverin in Ca2+-dependent membrane interaction. Science 259, 829–832. - PubMed
-
- Ellard-Ivey, M., Hopkins, R.B., White, T.J., and Lomax, T.L. (1999). Cloning, expression and N-terminal myristoylation of CpCPK1, a calcium-dependent protein kinase from zucchini (Cucurbita pepo L.). Plant Mol. Biol. 39, 199–208. - PubMed
-
- Galbiati, F., Guzzi, F., Magee, A.I., Milligan, G., and Parenti, M. (1996). Chemical inhibition of myristoylation of the G-protein Gil alpha by 2-hydroxymyristate does not interfere with its palmitoylation or membrane association: Evidence that palmitoylation, but not myristoylation, regulates membrane attachment. Biochem. J. 313, 717–720. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
