

Identification and characterization of a novel microtubule-based motor associated with membranous organelles in tobacco pollen tubes
- PMID: 11006343
- PMCID: PMC149081
- DOI: 10.1105/tpc.12.9.1719
Identification and characterization of a novel microtubule-based motor associated with membranous organelles in tobacco pollen tubes
Abstract
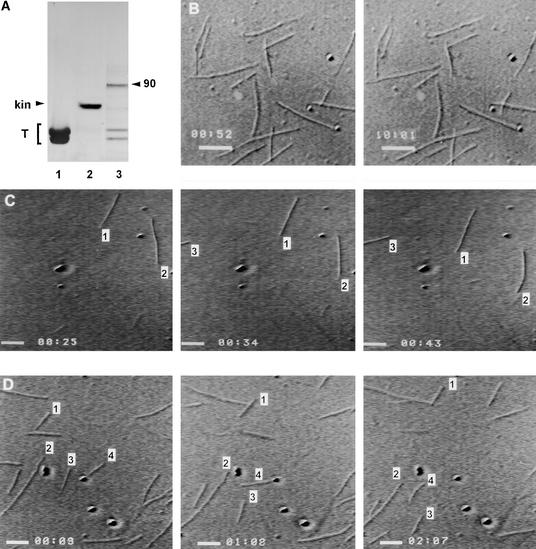
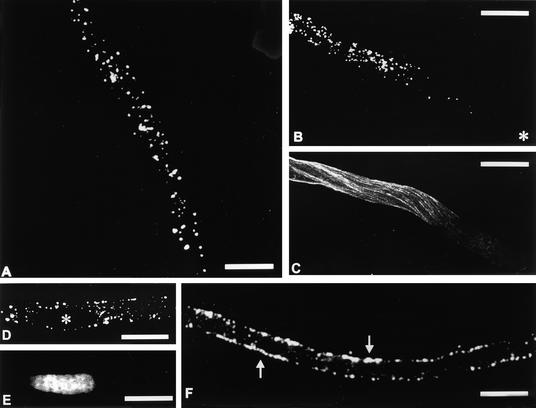
Pollen tube growth depends on the differential distribution of organelles and vesicles along the tube. The role of microtubules in organelle movement is uncertain, mainly because information at the molecular level is limited. In an effort to understand the molecular basis of microtubule-based movement, we isolated from tobacco pollen tubes polypeptides that cosediment with microtubules in an ATP-dependent manner. Major polypeptides released from microtubules by ATP (ATP-MAPs) had molecular masses of 90, 80, and 41 kD. Several findings indicate that the 90-kD ATP-MAP is a kinesin-related motor: binding of the polypeptide to microtubules was enhanced by the nonhydrolyzable ATP analog AMP-PNP; the 90-kD polypeptide reacted specifically with a peptide antibody directed against a highly conserved region in the motor domain of the kinesin superfamily; purified 90-kD ATP-MAP induced microtubules to glide in motility assays in vitro; and the 90-kD ATP-MAP cofractionated with microtubule-activated ATPase activity. Immunolocalization studies indicated that the 90-kD ATP-MAP binds to organelles associated with microtubules in the cortical region of the pollen tube. These findings suggest that the 90-kD ATP-MAP is a kinesin-related microtubule motor that moves organelles in the cortex of growing pollen tubes.
Figures








References
-
- Asada, T., and Collings, D. (1997). Molecular motors in higher plants. Trends Plant Sci. 2, 29–37.
-
- Asada, T., and Shibaoka, H. (1994). Isolation of polypeptides with microtubule-translocating activity from phragmoplasts of tobacco BY-2 cells. J. Cell Sci. 107, 2249–2257. - PubMed
-
- Asada, T., Kuriyama, R., and Shibaoka, H. (1997). TKRP125, a kinesin-related protein involved in the centrosome-independent organization of the cytokinetic apparatus in tobacco BY-2 cells. J. Cell Sci. 110, 179–189. - PubMed
-
- Åström, H., Sorri, O., and Raudaskoski, M. (1995). Role of microtubules in the movement of the vegetative nucleus and generative cell in tobacco pollen tubes. Sex. Plant Reprod. 8, 61–69.
-
- Baxter-Gabbard, K.L. (1972). A simple method for the large-scale preparation of sucrose gradients. FEBS Lett. 20, 117–119. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
