

Biogenesis of the chloroplast-encoded D1 protein: regulation of translation elongation, insertion, and assembly into photosystem II
- PMID: 11006346
- PMCID: PMC149084
- DOI: 10.1105/tpc.12.9.1769
Biogenesis of the chloroplast-encoded D1 protein: regulation of translation elongation, insertion, and assembly into photosystem II
Abstract
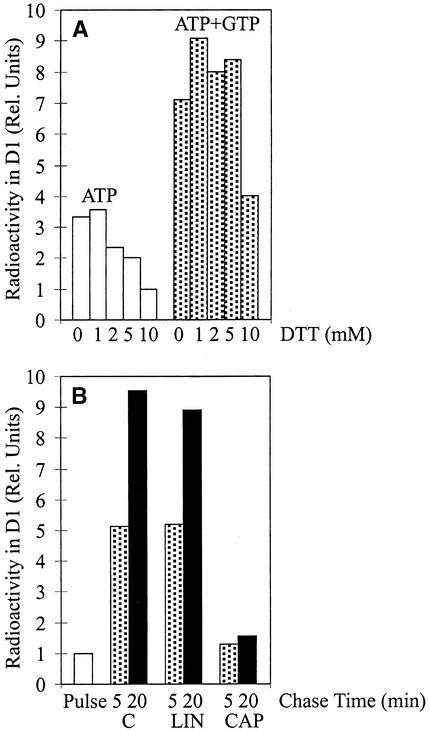
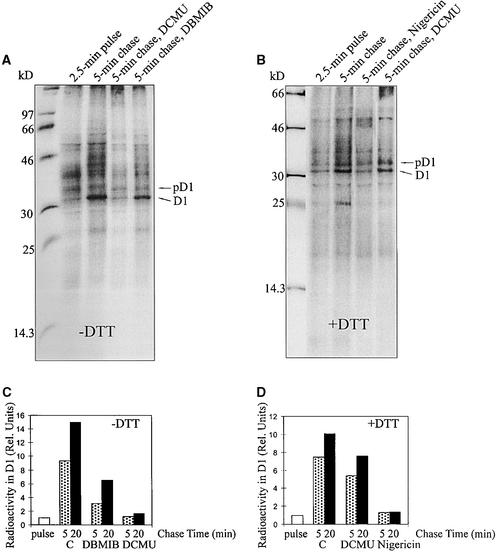
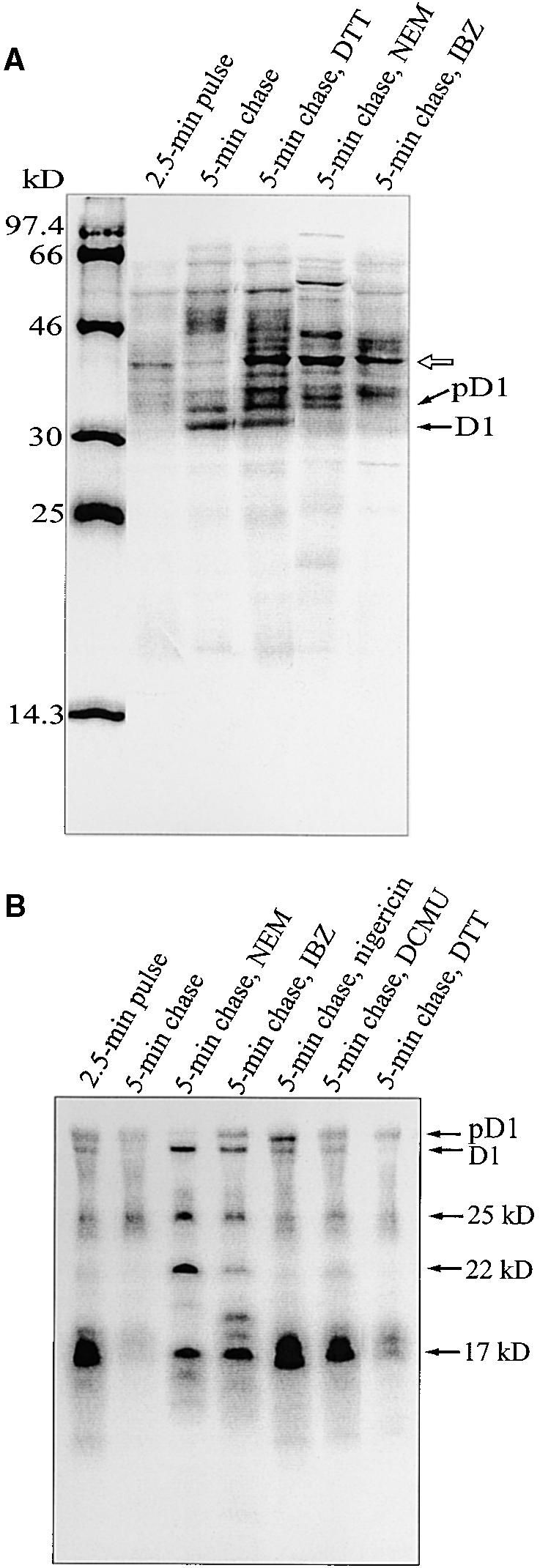
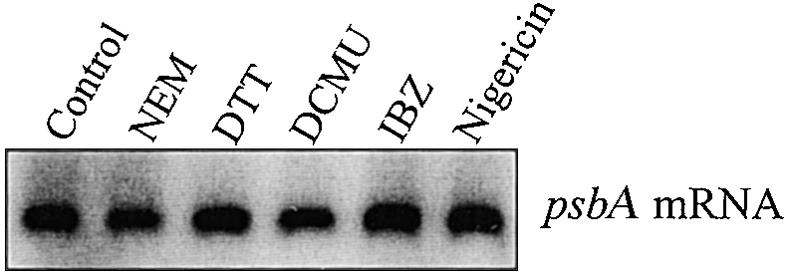
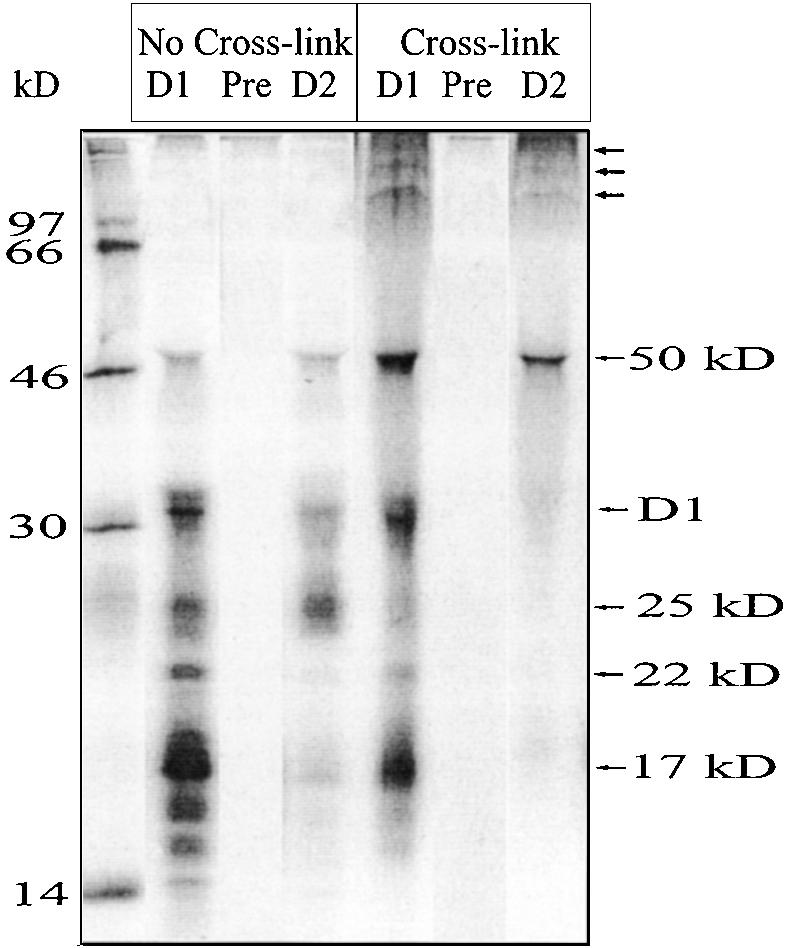
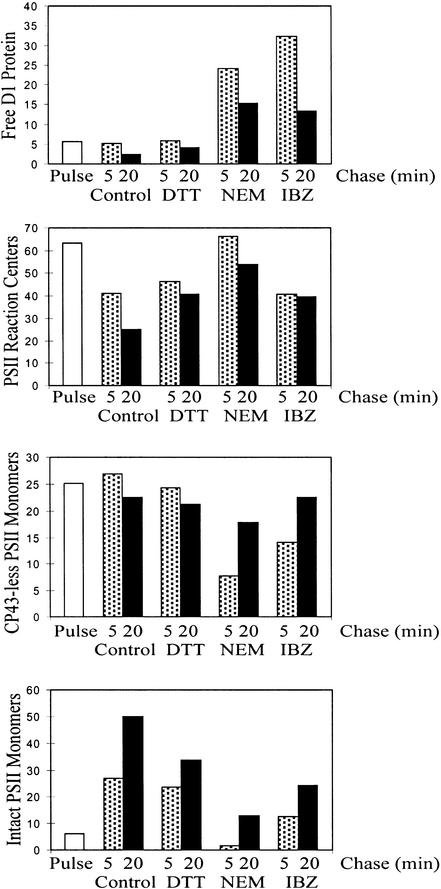
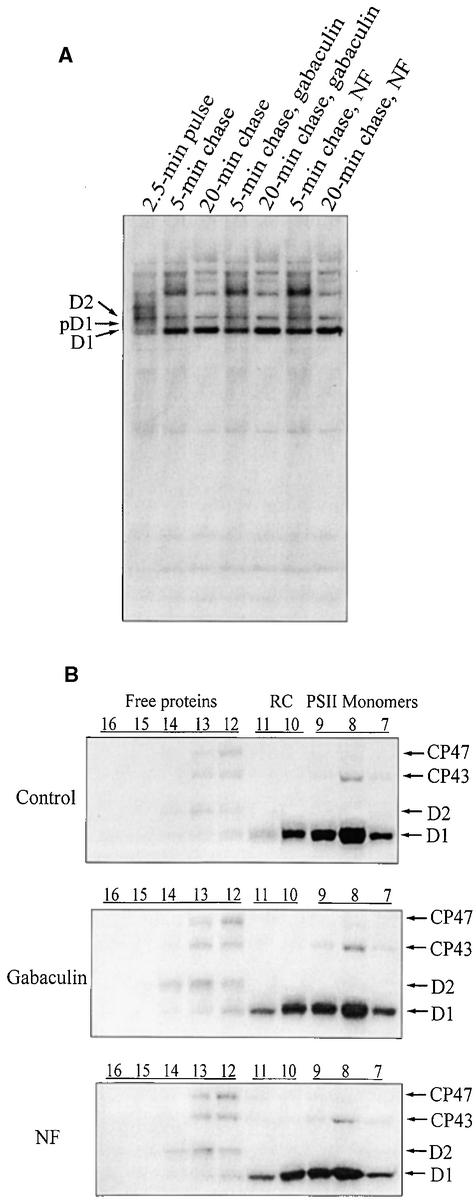
Regulation of translation elongation, membrane insertion, and assembly of the chloroplast-encoded D1 protein of photosystem II (PSII) was studied using a chloroplast translation system in organello. Translation elongation of D1 protein was found to be regulated by (1) a redox component that can be activated not only by photosynthetic electron transfer but also by reduction with DTT; (2) the trans-thylakoid proton gradient, which is absolutely required for elongation of D1 nascent chains on the thylakoid membrane; and (3) the thiol reactants N-ethylmaleimide (NEM) and iodosobenzoic acid (IBZ), which inhibit translation elongation with concomitant accumulation of distinct D1 pausing intermediates. These results demonstrate that D1 translation elongation and membrane insertion are tightly coupled and highly regulated processes in that proper insertion is a prerequisite for translation elongation of D1. Cotranslational and post-translational assembly steps of D1 into PSII reaction center and core complexes occurred independently of photosynthetic electron transfer or trans-thylakoid proton gradient but were strongly affected by the thiol reactants DTT, NEM, and IBZ. These compounds reduced the stability of the early PSII assembly intermediates, hampered the C-terminal processing of the precursor of D1, and prevented the post-translational reassociation of CP43, indicating a strong dependence of the D1 assembly steps on proper redox conditions and the formation of disulfide bonds.
Figures

References
-
- Adir, N., Shochat, S., and Ohad, I. (1990). Light-dependent D1 protein synthesis and translocation is regulated by reaction center II: Reaction center II serves as an acceptor for the D1 precursor. J. Biol. Chem. 265, 12563–12568. - PubMed
-
- Aro, E.-M., Virgin, I., and Andersson, B. (1993). Photoinhibition of photosystem II. Inactivation, protein damage and turnover. Biochim. Biophys. Acta 1143, 113–143. - PubMed
-
- Baena-Gonzalez, E., Barbato, R., and Aro, E.-M. (1999). Role of phosphorylation in the repair cycle and oligomeric structure of photosystem II. Planta 208, 196–204.
-
- Bowyer, J.R., Packer, J.C.L., McCormack, B.A., Whitelegge, J.P., Robinson, C., and Tayor, M. (1992). Carboxyl-terminal processing of the D1 protein and photoactivation of water splitting in photosystem II. J. Biol. Chem. 267, 5424–5433. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
