

Differential electron flow around photosystem I by two C(4)-photosynthetic-cell-specific ferredoxins
- PMID: 11013207
- PMCID: PMC302093
- DOI: 10.1038/sj.emboj.7593319
Differential electron flow around photosystem I by two C(4)-photosynthetic-cell-specific ferredoxins
Abstract
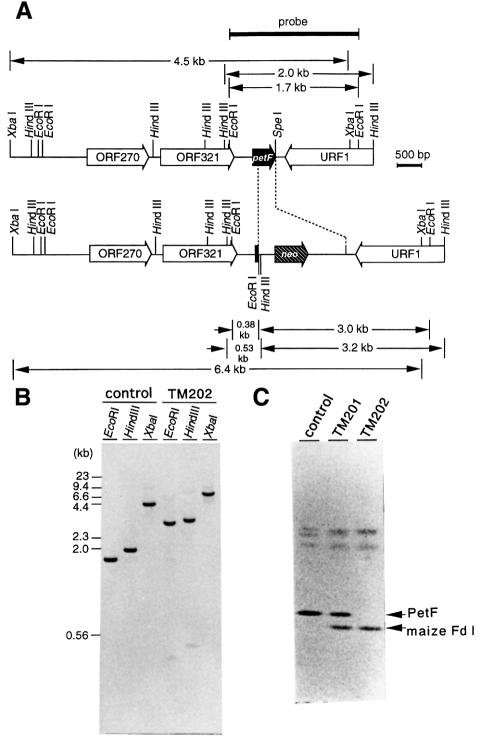
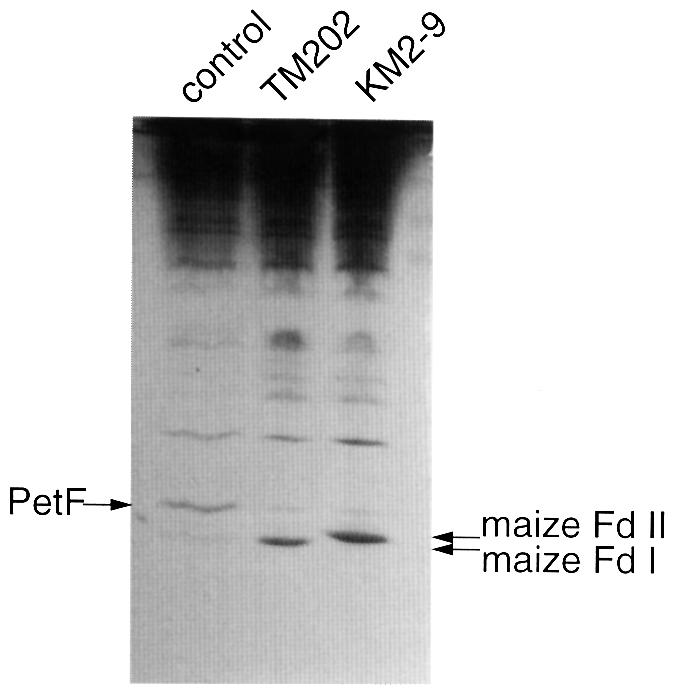
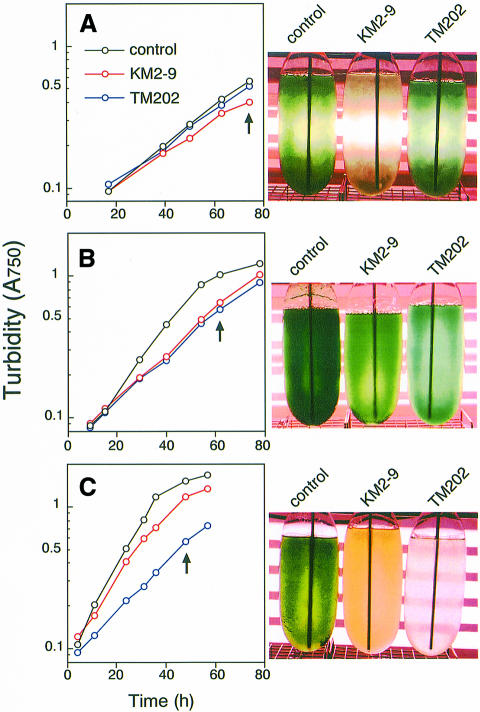
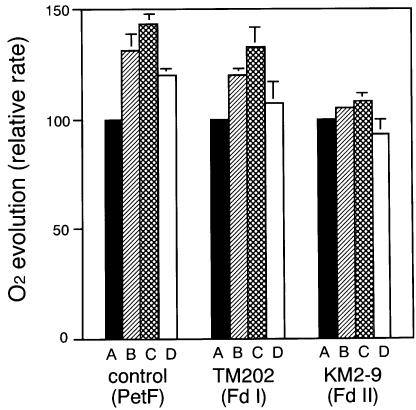
In the C(4) plant maize (Zea mays L.), two ferredoxin isoproteins, Fd I and Fd II, are expressed specifically in mesophyll and bundle-sheath cells, respectively. cDNAs for these ferredoxins were introduced separately into the cyanobacterium Plectonema boryanum with a disrupted endogenous ferredoxin gene, yielding TM202 and KM2-9 strains expressing Fd I and Fd II. The growth of TM202 was retarded under high light (130 micromol/m(2)/s), whereas KM2-9 grew at a normal rate but exhibited a nitrogen-deficient phenotype. Measurement of photosynthetic O(2) evolution revealed that the reducing power was not efficiently partitioned into nitrogen assimilation in KM2-9. After starvation of the cells in darkness, the P700 oxidation level under far-red illumination increased significantly in TM202. However, it remained low in KM2-9, indicating an active cyclic electron flow. In accordance with this, the cellular ratio of ATP/ADP increased and that of NADPH/NADP(+) decreased in KM2-9 as compared with TM202. These results demonstrated that the two cell type-specific ferredoxins differentially modulate electron flow around photosystem I.
Figures




References
-
- Bendall D.S. and Manasse,R.S. (1995) Cyclic photophosphorylation and electron transport. Biochim. Biophys. Acta, 1229, 23–38.
-
- Berger S., Ellersiek,U., Westhoff,P. and Steinmüller,K. (1993) Studies on the expression of NDH-H, a subunit of the HAD(P)H-plastoquinone-oxidoreductase of higher-plant chloroplasts. Planta, 190, 25–31.
-
- Canaani O. (1990) The role of cyclic electron flow around photosystem I and excitation energy distribution between the photosystems upon acclimation to high ionic strength in Dunaliella.Photochem. Photobiol., 52, 591–599.
-
- Cassing A., Boehme,H. and Schrautemeier,B. (1995) Nucleotide sequence, promoter structure, and expression of the petF1 gene (accession No. U33848) encoding the [2Fe–2S] Fd I from the nitrogen-fixing nonheterocystous cyanobacterium Plectonema boryanum PCC 73110. Plant Physiol., 109, 1499.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
