

Transfer of palmitate from phospholipids to lipid A in outer membranes of gram-negative bacteria
- PMID: 11013210
- PMCID: PMC302101
- DOI: 10.1093/emboj/19.19.5071
Transfer of palmitate from phospholipids to lipid A in outer membranes of gram-negative bacteria
Abstract
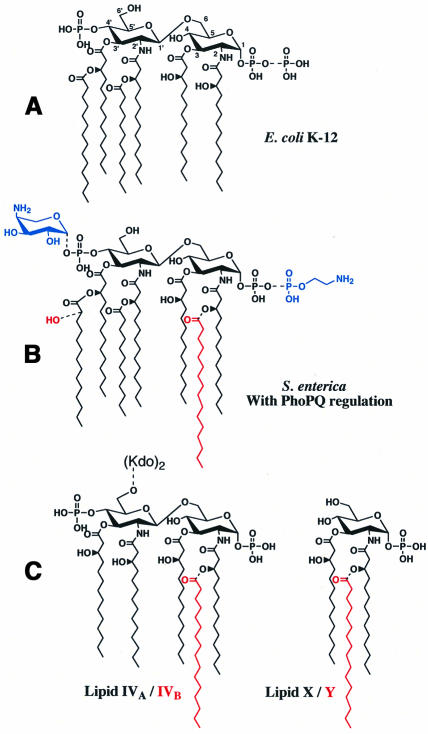
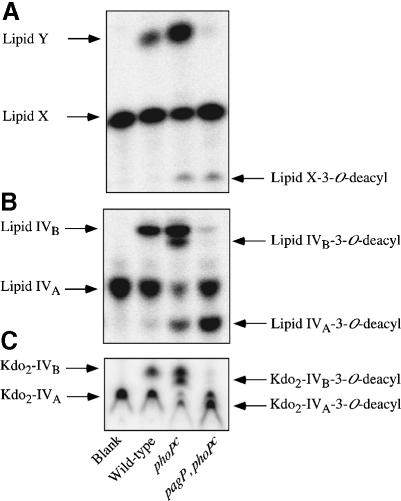
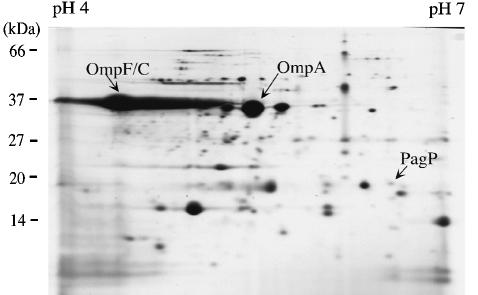
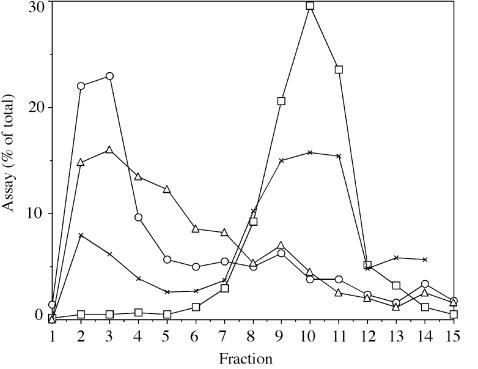
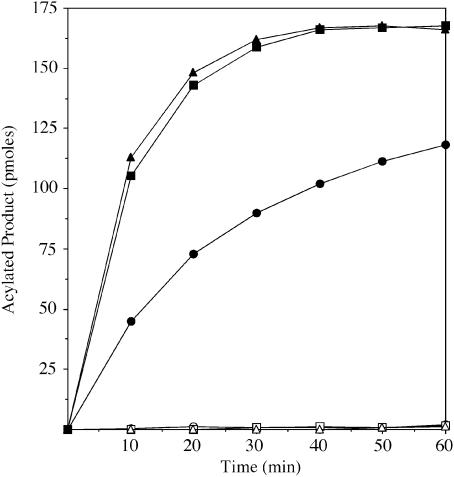
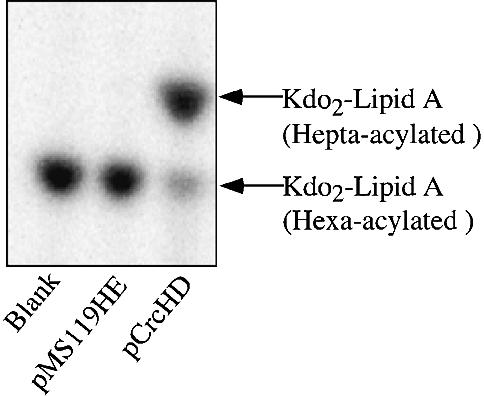
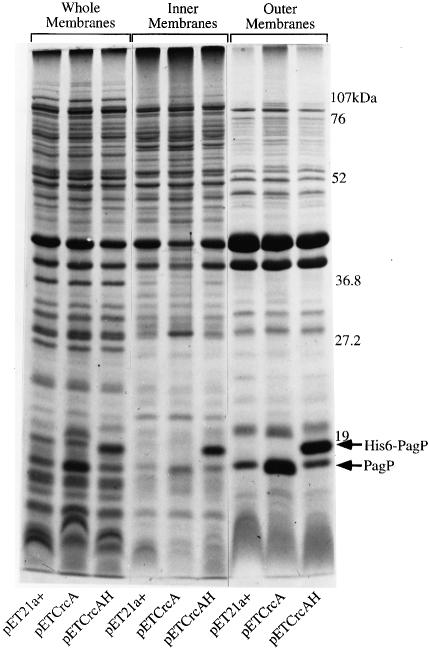
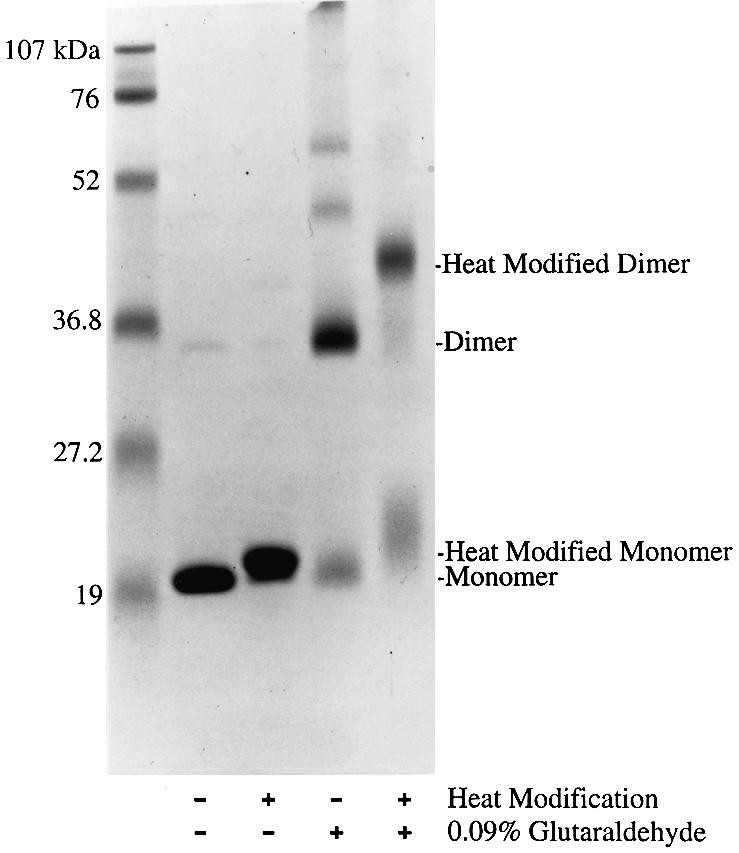
Regulated covalent modifications of lipid A are implicated in virulence of pathogenic Gram-negative bacteria. The Salmonella typhimurium PhoP/PhoQ-activated gene pagP is required both for biosynthesis of hepta-acylated lipid A species containing palmitate and for resistance to cationic anti-microbial peptides. Palmitoylated lipid A can also function as an endotoxin antagonist. We now show that pagP and its Escherichia coli homolog (crcA) encode an unusual enzyme of lipid A biosynthesis localized in the outer membrane. PagP transfers a palmitate residue from the sn-1 position of a phospholipid to the N-linked hydroxymyristate on the proximal unit of lipid A (or its precursors). PagP bearing a C-terminal His(6)-tag accumulated in outer membranes during overproduction, was purified with full activity and was shown by cross-linking to behave as a homodimer. PagP is the first example of an outer membrane enzyme involved in lipid A biosynthesis. Additional pagP homologs are encoded in the genomes of Yersinia and Bordetella species. PagP may provide an adaptive response toward both Mg(2+) limitation and host innate immune defenses.
Figures
References
-
- Basu S.S., York,J.D. and Raetz,C.R.H. (1999) A phosphotransferase that generates phosphatidylinositol 4-phosphate (PtdIns-4-P) from phosphatidylinositol and lipid A in Rhizobium leguminosarum. A membrane-bound enzyme linking lipid A and PtdIns-4-P biosynthesis. J. Biol. Chem., 274, 11139–11149. - PMC - PubMed
-
- Brabetz W., Muller-Loennies,S., Holst,O. and Brade,H. (1997) Deletion of the heptosyltransferase genes rfaC and rfaF in Escherichia coli K-12 results in an Re-type lipopolysaccharide with a high degree of 2-aminoethanol phosphate substitution. Eur. J. Biochem., 247, 716–724. - PubMed
-
- Brozek K.A. and Raetz,C.R.H (1990) Biosynthesis of lipid A in Escherichia coli. Acyl carrier protein-dependent incorporation of laurate and myristate. J. Biol. Chem., 265, 15410–15417. - PubMed
-
- Brozek K.A., Bulawa,C.E. and Raetz,C.R.H. (1987) Biosynthesis of lipid A precursors in Escherichia coli. A membrane-bound enzyme that transfers a palmitoyl residue from a glycerophospholipid to lipid X. J. Biol. Chem., 262, 5170–5179. - PubMed
-
- Brozek K.A., Hosaka,K., Robertson,A.D. and Raetz,C.R.H. (1989) Biosynthesis of lipopolysaccharide in Escherichia coli. Cytoplasmic enzymes that attach 3-deoxy-d-manno-octulosonic acid to lipid A. J. Biol. Chem., 264, 6956–6966. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
