

doi: 10.1093/emboj/19.19.5202.
Targeting the chromatin-remodeling MSL complex of Drosophila to its sites of action on the X chromosome requires both acetyl transferase and ATPase activities
Affiliations
- PMID: 11013222
- PMCID: PMC302094
- DOI: 10.1093/emboj/19.19.5202
Item in Clipboard
Targeting the chromatin-remodeling MSL complex of Drosophila to its sites of action on the X chromosome requires both acetyl transferase and ATPase activities
EMBO J.
.
Abstract
Dosage compensation in Drosophila is mediated by a multiprotein, RNA-containing complex that associates with the X chromosome at multiple sites. We have investigated the role that the enzymatic activities of two complex components, the histone acetyltransferase activity of MOF and the ATPase activity of MLE, may have in the targeting and association of the complex with the X chromosome. Here we report that MLE and MOF activities are necessary for complexes to access the various X chromosome sites. The role that histone H4 acetylation plays in this process is supported by our observations that MOF overexpression leads to the ectopic association of the complex with autosomal sites.
Figures
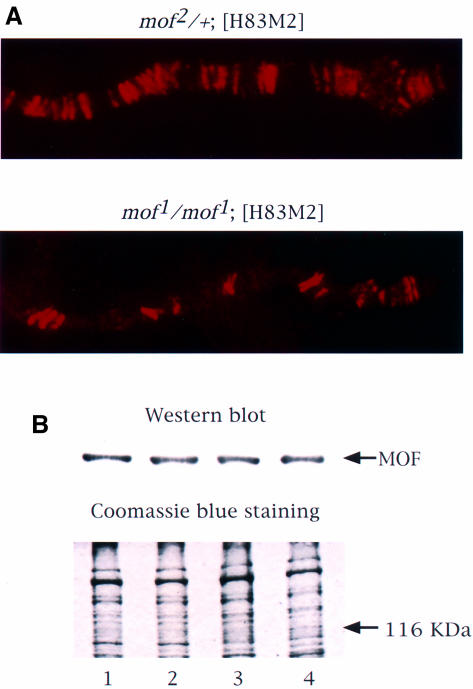
Fig. 1. MSL complexes are targeted only to the entry sites on the X chromosome in the presence of mof1. (A) Polytene chromosomes from control mof2/+; [H83M2] and mof1/mof1; [H83M2] female larvae were immunolabeled with antiserum against MOF. The distribution of the MOF protein is the same as that observed for all of the other MSLs (Gu et al., 1998). (B) MOF is expressed at a similar level in wild-type males (lane 1), wild-type females (lane 2), mof1/mof1; [H83M2] females (lane 3) and mof2/+; [H83M2] females (lane 4). Protein extracts prepared from adult fly heads were resolved by 7.5% SDS–PAGE. The proteins were transferred to nitrocellulose membrane and incubated with anti-MOF serum. A parallel gel was stained with Coomassie Brilliant Blue R-250 to monitor the amount of protein loaded.
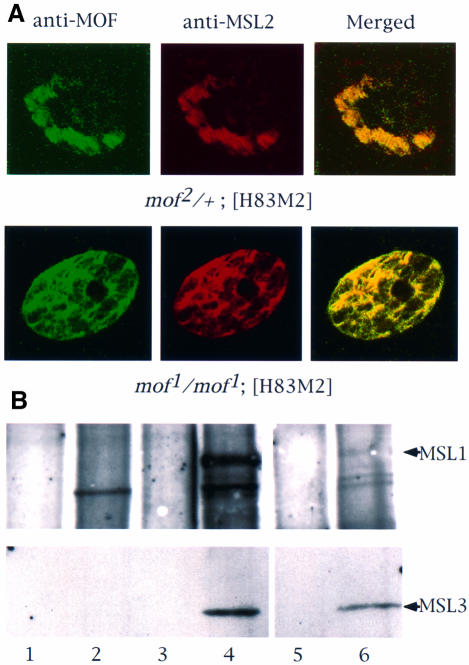
Fig. 2. In the absence of MOF activity, MSL complexes are found in the nucleoplasm. (A) Whole salivary glands from mof1/mof1; [H83M2] and mof2/+; [H83M2] larvae were stained for MSL2 and MOF by indirect immunofluorescent labeling. In the nuclei of larvae expressing mof+, the two antigens are co-localized along the paired X chromo somes. In mof1 homozygotes, the MSL complex appears in the interchromosomal spaces. Note that it must also be present at the entry sites on the X chromosomes although these cannot be resolved in this material. (B) Co-immunoprecipitation of MSL1 and MSL3. Protein extracts prepared from wild-type female (lanes 1 and 2), wild-type male (lanes 3 and 4) and mof1/mof1; [H83M2] female (lanes 5 and 6) flies were immunoprecipitated with pre-immune (lanes 1, 3 and 5) or MSL1 antisera (lanes 2, 4 and 6). The precipitate was analyzed by western blotting using MSL1 and MSL3 antisera. (C) Localization of roX1 RNA, determined by in situ hybridization in whole salivary glands. The RNA is localized to a limited area corresponding to the X chromosome in male nuclei and is absent in the nuclei of wild-type females. In mof1/mof1; [H83M2] females, roX1 RNA is dispersed in a pattern that mimics that of the MSLs. The arrows mark the nuclear envelope. (D) In situ hybridization of roX1 RNA on the polytene chromosomes of mof1/mof1; [H83M2] female larvae. (a) Propidium iodide staining of a nucleus; (b) roX1 is present at the entry sites on the X chromosome in the same nucleus; (c) a stretched region of X chromosome from another nucleus.
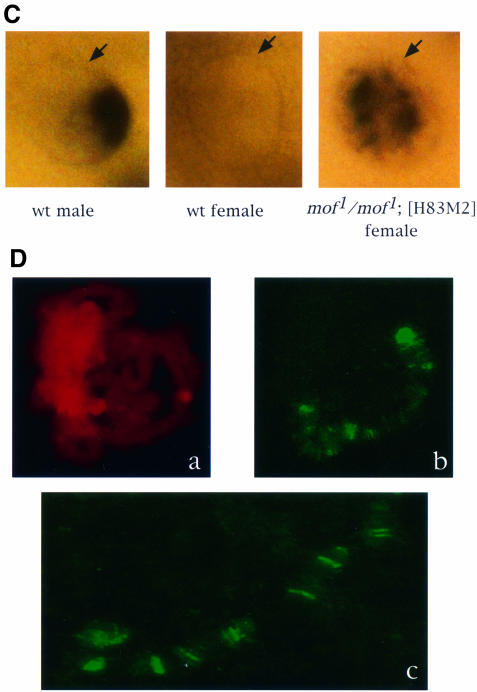
Fig. 2. In the absence of MOF activity, MSL complexes are found in the nucleoplasm. (A) Whole salivary glands from mof1/mof1; [H83M2] and mof2/+; [H83M2] larvae were stained for MSL2 and MOF by indirect immunofluorescent labeling. In the nuclei of larvae expressing mof+, the two antigens are co-localized along the paired X chromo somes. In mof1 homozygotes, the MSL complex appears in the interchromosomal spaces. Note that it must also be present at the entry sites on the X chromosomes although these cannot be resolved in this material. (B) Co-immunoprecipitation of MSL1 and MSL3. Protein extracts prepared from wild-type female (lanes 1 and 2), wild-type male (lanes 3 and 4) and mof1/mof1; [H83M2] female (lanes 5 and 6) flies were immunoprecipitated with pre-immune (lanes 1, 3 and 5) or MSL1 antisera (lanes 2, 4 and 6). The precipitate was analyzed by western blotting using MSL1 and MSL3 antisera. (C) Localization of roX1 RNA, determined by in situ hybridization in whole salivary glands. The RNA is localized to a limited area corresponding to the X chromosome in male nuclei and is absent in the nuclei of wild-type females. In mof1/mof1; [H83M2] females, roX1 RNA is dispersed in a pattern that mimics that of the MSLs. The arrows mark the nuclear envelope. (D) In situ hybridization of roX1 RNA on the polytene chromosomes of mof1/mof1; [H83M2] female larvae. (a) Propidium iodide staining of a nucleus; (b) roX1 is present at the entry sites on the X chromosome in the same nucleus; (c) a stretched region of X chromosome from another nucleus.
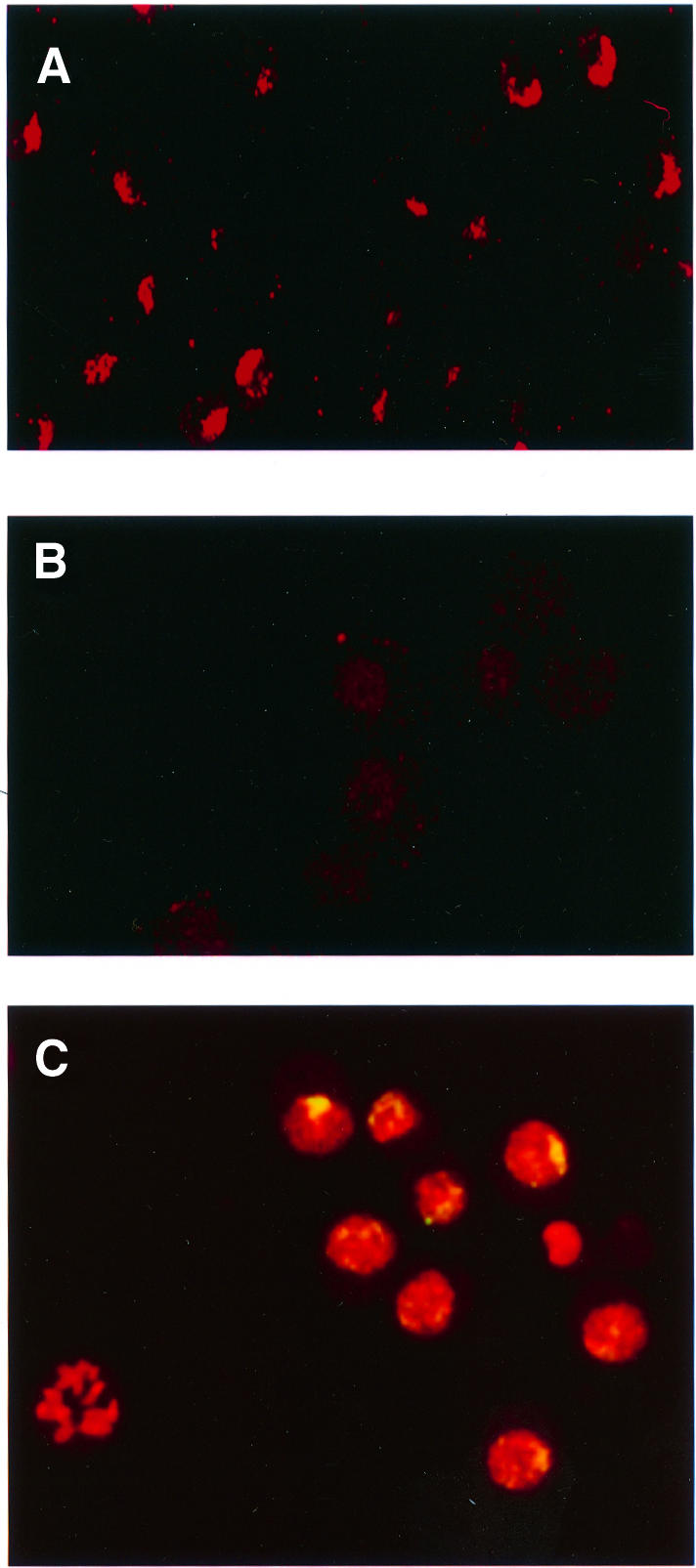
Fig. 3. Overexpression of MOF1 in S2 cells causes a reduction in the level of H4Ac16 present on the X chromosomes and relocation of MSL complexes. Histone H4Ac16 in S2 cells transfected with the vector (A) or with mof1 cDNA (B) and induced with CuSO4. (C) Merged image of induced S2-mof1 cells stained for MSL1 (green) and counterstained for nucleic acids with propidium iodide (red). The yellow color indicates the presence of the MSL1 protein.
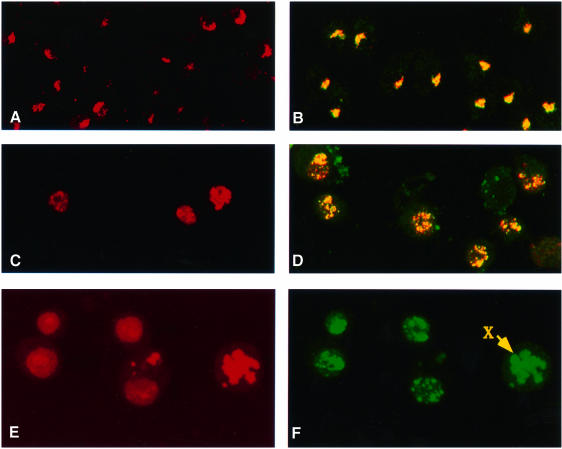
Fig. 4. MSL complexes are targeted to autosomal sites in S2 cells overexpressing MOF. (A) Histone H4Ac16 staining is restricted to the X chromosome in cells transfected with the vector induced with CuSO4. (B) Co-localization to the X chromosome is also seen in the merged image of immunofluorescent labeling for MSL1 (red) and MSL3 (green) in similar cells. (C) In contrast, H4Ac16 is widespread throughout the nuclei in cells transfected with mof cDNA and induced with CuSO4. (D) Merged images of immunofluorescent labeling for MSL1 (red) and MSL3 (green) in similar cells. The complex distribution generally follows the distribution of H4Ac16. (E) Induced S2-mof cells were stained for nucleic acids with propidium iodide. (F) The same cells were immunolabeled with anti-MSL1 serum. The distribution of MSL1 and, by inference, of the MSL complex is clearly autosomal. The arrow points to a chromosome that is more intensely stained than the others and is presumably the X chromosome.
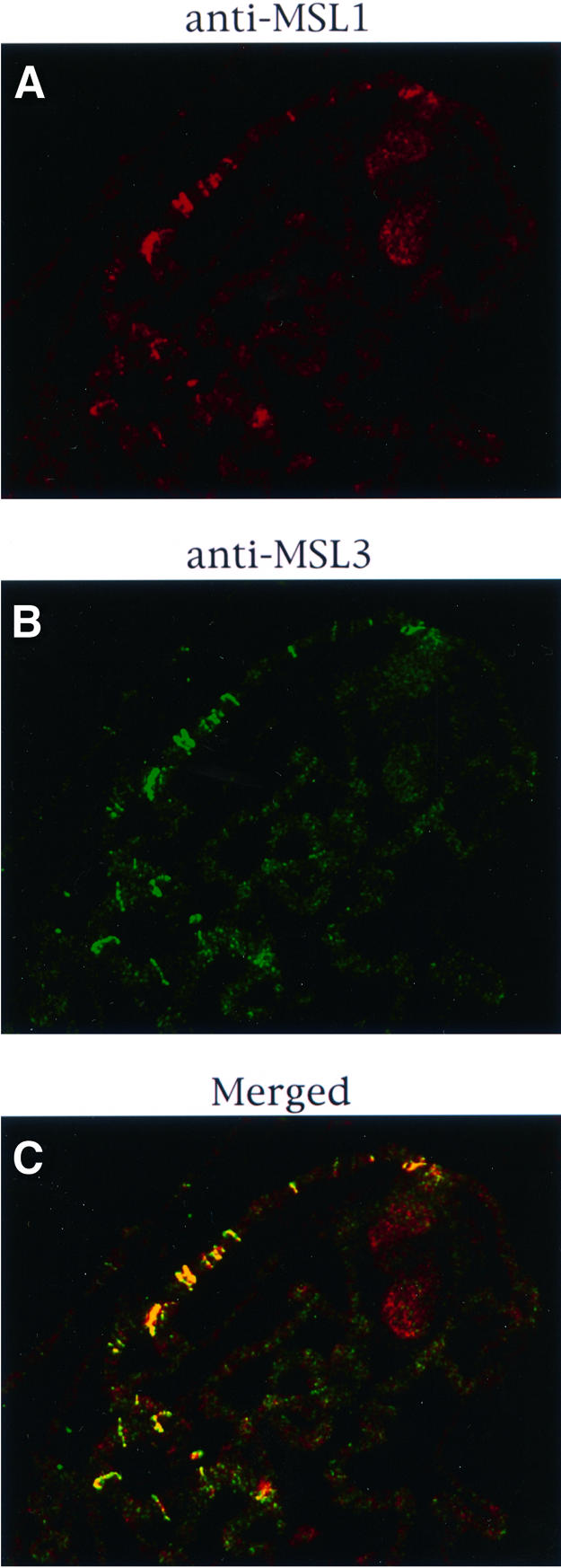
Fig. 5. Effect of the mleGET mutation on the localization of the MSL complex. Polytene chromosomes from mle1/mle1; [mle-GET]/[H83M2] larvae were stained for MSL1 (A) or MSL3 (B). As seen in the merged image (C), these two MSLs are co-localized at the entry sites.
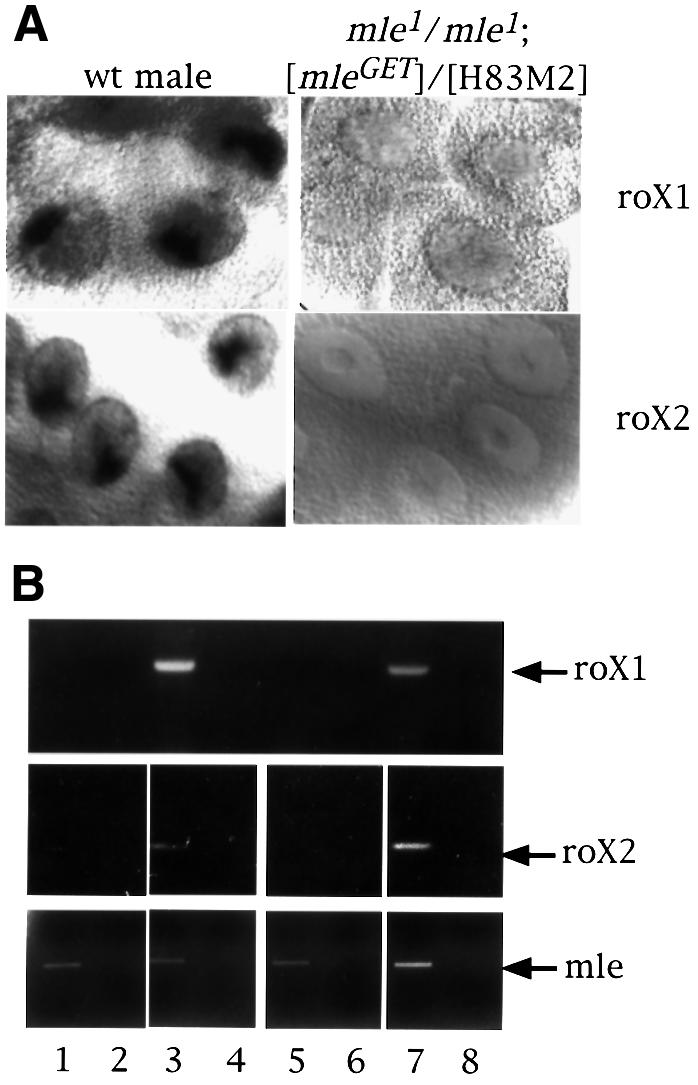
Fig. 6. roX RNA is absent in females that produce only the MLEGET mutant protein. (A) Localization of roX1 and roX2 RNA by in situ hybridization in whole salivary glands from a wild-type male and a mle1/mle1; [mle-GET]/[H83M2] female larva. (B) RT–PCR for roX RNA. Total RNA from adult flies of wild-type females (lanes 1 and 2), wild-type males (lanes 3 and 4), mle1/mle1; [mle-GET]/[H83M2] females (lanes 5 and 6) and mle1/+; [mle-GET]/[H83M2] females (lanes 7 and 8) were reverse transcribed (lanes 1, 3, 5 and 7) or not (lanes 2, 4, 6 and 8) and then amplified for roX1 or roX2 RNA by PCR. The lower panel shows the amplification of MLE RNA by RT–PCR in the same samples.
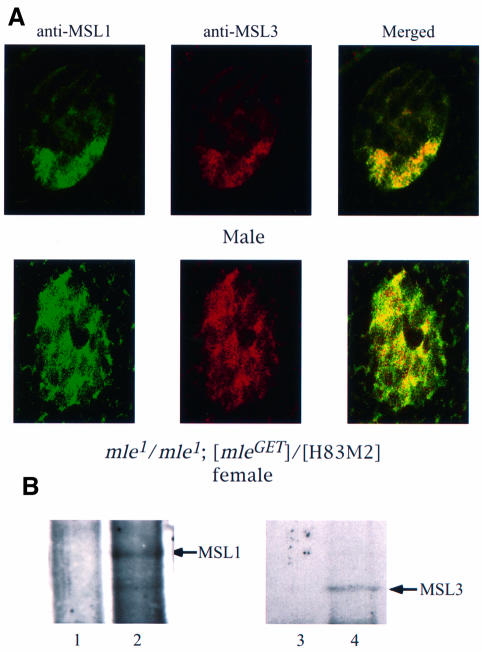
Fig. 7. In the absence of MLE ATPase activity, MSL complexes lacking roX RNA are found in the nucleoplasm. (A) Whole salivary glands from mle1/mle1; [mle-GET] and wild-type male larvae were stained for MSL1 and MSL3 by indirect immunofluorescent labeling. In the nuclei of male larvae expressing only mleGET, the two antigens are co-localized along the X chromosome. In the nuclei of female larvae from mle1/mle1; [mle-GET], although some MSL1 (green) and MSL3 (red) proteins appear to be independent of each other, a substantial amount of these proteins are associated (yellow, in the merged image), leading to the conclusion that the MSL complex exists in the interchromosomal spaces. Note that, as stated in the legend of Figure 5, the complex must also be present at the entry sites on the X chromosomes, although these cannot be resolved in this material. (B) Co-immunoprecipitation of MSL1 and MSL3. Protein extracts prepared from mle1/mle1; [mle-GET] female flies were immuno precipitated with pre-immune (lanes 1 and 3) or MSL1 antisera (lanes 2 and 4). The precipitate was analyzed by western blotting using MSL1 and MSL3 antisera. Note that the lanes presented in this figure were obtained from the gel illustrated in Figure 2, which includes the wild-type male and female controls.
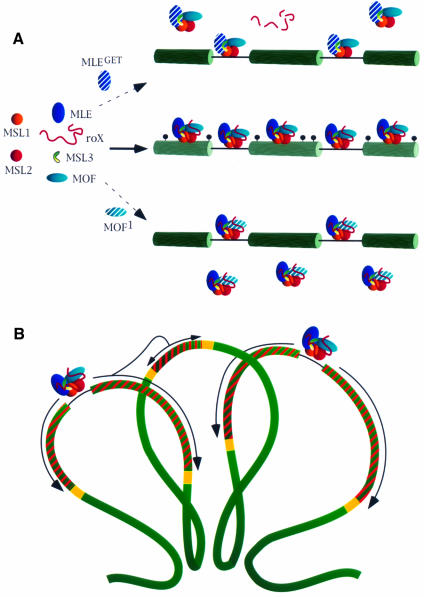
Fig. 8. A model for the association and spreading of the MSL complex along the X chromosome. (A) In wild type, the MSL complex assembles at the roX-bearing entry sites, moves to other sites and acetylates histone H4 of neighboring nucleosomes at Lys16 (indicated as black circles). In the absence of the ATPase activity of MLE (MLEGET), the association of the complex with roX RNA is unstable. Free complexes devoid of roX RNA and unable to access the X chromosome are present in the nucleoplasm. If the acetylase function of MOF is impaired by mutation (MOF1), complexes are formed; although they can access other entry sites, they cannot spread along the X chromosome and are found, unbound, in the nucleoplasm. (B) Wild-type MSL complexes present at the entry sites spread along the X chromosome by acetylating nucleosomes. Spreading of the complexes and concomitant acetylation (indicated by the striped regions) may be interrupted by some type of boundary elements (yellow boxes). The looping of the chromatid fiber into rosettes may enable spreading of the complex from a region where acetylation initiated at an entry site to a region where no entry sites are present (from the left to the central loop in the diagram).
Similar articles
-
The roX genes encode redundant male-specific lethal transcripts required for targeting of the MSL complex.EMBO J. 2002 Mar 1;21(5):1084-91. doi: 10.1093/emboj/21.5.1084. EMBO J. 2002. PMID: 11867536 Free PMC article.
-
Targeting of MOF, a putative histone acetyl transferase, to the X chromosome of Drosophila melanogaster.Dev Genet. 1998;22(1):56-64. doi: 10.1002/(SICI)1520-6408(1998)22:1<56::AID-DVG6>3.0.CO;2-6. Dev Genet. 1998. PMID: 9499580
-
Sex-biased transcription enhancement by a 5' tethered Gal4-MOF histone acetyltransferase fusion protein in Drosophila.BMC Mol Biol. 2010 Nov 9;11:80. doi: 10.1186/1471-2199-11-80. BMC Mol Biol. 2010. PMID: 21062452 Free PMC article.
-
The right dose for every sex.Chromosoma. 2007 Apr;116(2):95-106. doi: 10.1007/s00412-006-0089-x. Epub 2006 Nov 24. Chromosoma. 2007. PMID: 17124606 Free PMC article. Review.
-
Chromatin mechanisms in Drosophila dosage compensation.Prog Mol Subcell Biol. 2005;38:123-49. doi: 10.1007/3-540-27310-7_5. Prog Mol Subcell Biol. 2005. PMID: 15881893 Review.
Cited by
-
Zinc finger protein Zn72D promotes productive splicing of the maleless transcript.Mol Cell Biol. 2007 Dec;27(24):8760-9. doi: 10.1128/MCB.01415-07. Epub 2007 Oct 8. Mol Cell Biol. 2007. PMID: 17923683 Free PMC article.
-
The MLE subunit of the Drosophila MSL complex uses its ATPase activity for dosage compensation and its helicase activity for targeting.Mol Cell Biol. 2008 Feb;28(3):958-66. doi: 10.1128/MCB.00995-07. Epub 2007 Nov 26. Mol Cell Biol. 2008. PMID: 18039854 Free PMC article.
-
The roX genes encode redundant male-specific lethal transcripts required for targeting of the MSL complex.EMBO J. 2002 Mar 1;21(5):1084-91. doi: 10.1093/emboj/21.5.1084. EMBO J. 2002. PMID: 11867536 Free PMC article.
-
The Drosophila roX1 RNA gene can overcome silent chromatin by recruiting the male-specific lethal dosage compensation complex.Genetics. 2003 Jun;164(2):565-74. doi: 10.1093/genetics/164.2.565. Genetics. 2003. PMID: 12807777 Free PMC article.
-
Dosage compensation in Drosophila.Cold Spring Harb Perspect Biol. 2015 May 1;7(5):a019398. doi: 10.1101/cshperspect.a019398. Cold Spring Harb Perspect Biol. 2015. PMID: 25934013 Free PMC article. Review.
References
-
- Akhtar A. and Becker,P.B. (2000) Activation of transcription through histone H4 acetylation by MOF, an acetyltransferase essential for dosage compensation in Drosophila. Mol. Cell, 5, 367–375. - PubMed
-
- Akhtar A., Zinc,D. and Becker,P.B. (2000) Chromodomains are protein–RNA interaction modules. Nature, in press. - PubMed
-
- Bashaw G.J. and Baker,B.S. (1995) The msl-2 dosage compensation gene of Drosophila encodes a putative DNA-binding protein whose expression is sex-specifically regulated by Sex-lethal. Development, 121, 3245–3258. - PubMed
-
- Bone J.R., Lavender,J., Richman,R., Palmer,M.J., Turner,B.M. and Kuroda,M.I. (1994) Acetylated histone H4 on the male X chromosome is associated with dosage compensation in Drosophila. Genes Dev., 8, 96–104. - PubMed
-
- Borrow J. et al. (1996) The translocation t(8;16)(p11;p13) of acute myeloid leukaemia fuses a putative acetyltransferase to the CREB-binding protein. Nature Genet., 14, 33–41. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
