

Increase of androgen-induced cell death and androgen receptor transactivation by BRCA1 in prostate cancer cells
- PMID: 11016951
- PMCID: PMC17187
- DOI: 10.1073/pnas.190353897
Increase of androgen-induced cell death and androgen receptor transactivation by BRCA1 in prostate cancer cells
Abstract
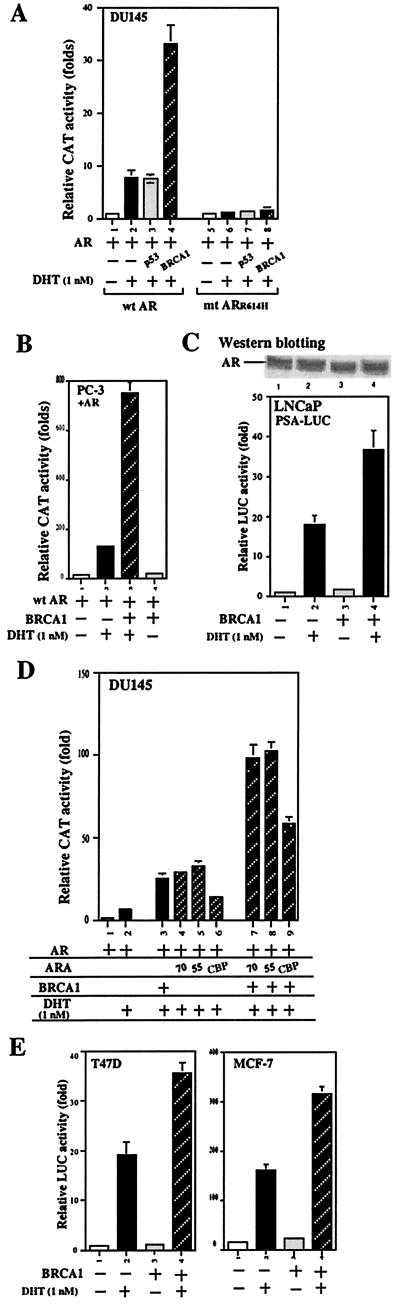
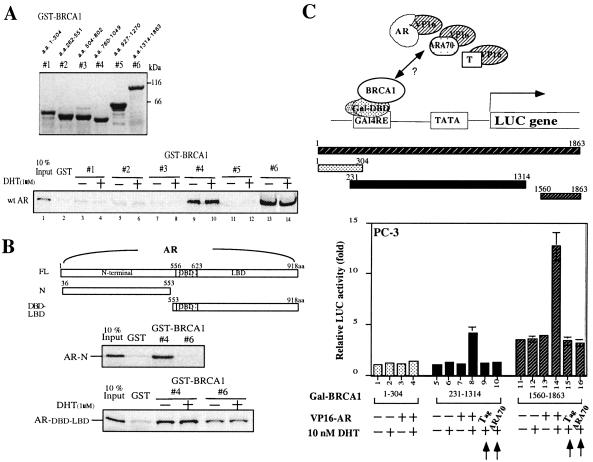
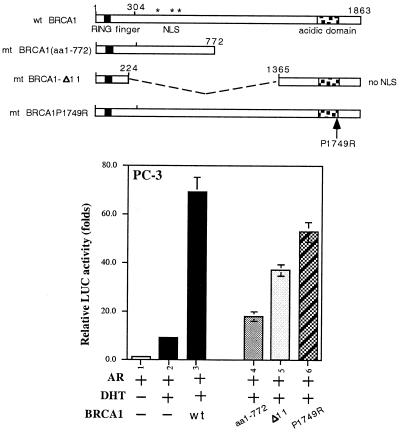
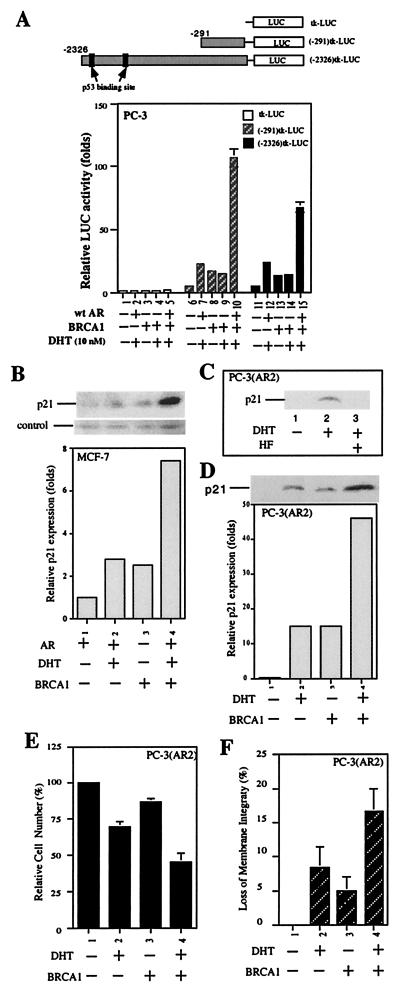
Although mutations of the breast cancer susceptibility gene 1 (BRCA1) may play important roles in breast and prostate cancers, the detailed mechanism linking the functions of BRCA1 to these two hormone-related tumors remains to be elucidated. Here, we report that BRCA1 interacts with androgen receptor (AR) and enhances AR target genes, such as p21((WAF1/CIP1)), that may result in the increase of androgen-induced cell death in prostate cancer cells. The BRCA1-enhanced AR transactivation can be further induced synergistically with AR coregulators, such as CBP, ARA55, and ARA70. Together, these data suggest that the BRCA1 may function as an AR coregulator and play positive roles in androgen-induced cell death in prostate cancer cells and other androgen/AR target organs.
Figures
References
-
- Futreal P A, Liu Q, Shattuck-Eidens D, Cochran C, Harshman K, Tavtigian S, Bennett L M, Haugen-Strano A, et al. Science. 1994;266:120–122. - PubMed
-
- Ford D, Easton D F, Bishop D T, Narod S A, Goldgar D E. Lancet. 1994;343:692–695. - PubMed
-
- Zhang H, Somasundaram K, Peng Y, Tian H, Bi H, Zhang D, Weber B L, El-Deiry W S. Oncogene. 1998;16:1713–1721. - PubMed
-
- Dawit T, Parvin H, Parvin J D. J Biol Chem. 1999;274:2113–2117. - PubMed
-
- Scully R, Chen J, Plug A, Xiao Y, Weaver D, Feunteun J, Ashley T, Livingston D M. Cell. 1997;88:265–275. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
